1.5 Microevolution, Macroevolution, and the Synthetic Theory
The first systematic attempt to categorize different levels of evolution was made by Richard B. Goldschmidt (1878-1958) (1). He took the original [49] allusion of Dobzhansky (2) to microevolution as the evolutionary process observable within man's lifetime and gave it an experimental meaning. Goldschmidt believed geneticists could use microevolution to analyze the variants of natural populations as well as observe evolutionary changes in controlled breeding studies in the laboratory. Macroevolution (megaevolution), a term coined by Simpson (3), was perceived as the territory of the paleontologist, the comparative anatomist, and the embryologist. Therefore, Goldschmidt interpreted microevolution as the observable changes that give rise to variations in experimental and natural populations within a species and macroevolution as the historical evolution of the "good" species that have been placed in the higher taxonomic categories. Furthermore, these macroevolutionary changes have been correlated with die geological time scale.
[50] Neo-Darwinists claimed that the accumulation of gene mutations and the isolation and selection of new variants that continue to undergo the same process accounted for all evolutionary diversification. Goldschmidt argued that these processes cannot he an all-encompassing mechanism and thus did not suffice for an understanding of macroevolution. He challenged the Neo-Darwinists to explain the evolution of 18 features by accumulation and selection of small mutations. The features included hair in mammals, feathers in birds, segmentation of arthropods and vertebrates, visceral arches, muscles, nerves, teeth, shells of mollusks, ectoskeletons, compound eyes, blood circulation, and alternation of generations. He proposed a novel concept of systemic mutation to account for macroevolution. This concept will be discussed in detail in 1.3.3.2.a.3. Neo-Darwinists, while unable to meet the challenges posed by
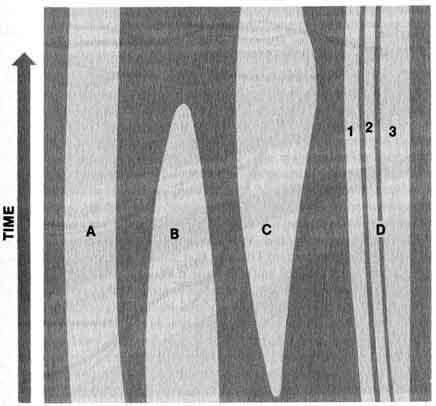
[51] Goldschmidt, found his alternative explanation untenable and adhered to their assertion that microevolution and macroevolution are not qualitatively distinct. They believed the accumulation of point-mutations by natural selection not only will lead to the development of new varieties within a species, but it can also account for the major features of macroevolution (3).Figure 1.8. An adaptive grid diagram of the evolution of terrestrial plants, indicating major breakthroughs and invasions of new adaptive zones. Reprinted, by permission. copyright 1969. Savage, J.M. Evolution. 2nd ed. New York, Holt, Rinehart, and Winston: 1969.
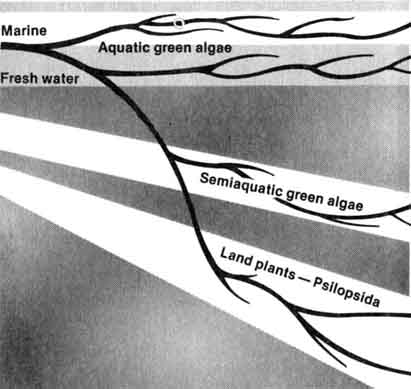
According to present-day Neo-Darwinists, the interaction of organisms and their environment produced a series of adaptive zones or fields (4). Each adaptive zone may also consist of subzones (Figure 1.7). For example all aquatic green algae occur in the aquatic adaptive zone, but the fresh water and the marine green algae are confined by their respective adaptive subzones. Microevolution involves the crossing of evolutionary lines within subdivisions of the subzones. However, macroevolution involves the crossing of evolutionary lines between one major zone or subzone into another. Evolution among green algae in the marine and flesh water subzones would be called microevolution, whereas macroevolution would involve major breakthroughs into new adaptive zones. A hypothetical macroevolutionary scheme constructed on the basis of geographical distribution of algae and land plants is represented in Figure 1.8.
1.5.1 Speciation. Neo-Darwinists account for the origin of species by the process of speciation. It is an extrapolation of variations seen in populations subjected to artificial selection. The major criterion that establishes a new species is the formation of reproductive isolation. Several of the mechanisms of reproductive isolation have been described earlier in this section. While Goldschmidt maintained that speciation occurs apart from the point mutation-selection scheme by a process of macrogenesis or saltation in which a "hopeful" monster arises by a rearrangement of the intrachromosomal pattern, this is rejected by the Neo-Darwinists. The mode of speciation advocated by Goldschmidt and others involves the establishment of reproductive isolation followed by development of geographical barriers. This view has been called sympatric speciation. It is a much refined form of the classical mutationist position that states that speciation takes place when reproductive isolation of a new population occurs in the presence of the parent species by disruptive selection, seasonal isolation, and polyploidy. In contrast, most Neo-Darwinists maintain that geographic speciation followed by reproductive isolation is the dominant evolutionary mode (5).
Neo-Darwinists have developed several patterns of evolution to account for trends in the evolutionary scheme constructed from the fossil record and the morphological differences observed in natural populations. [52]
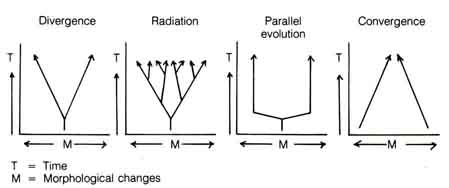
Rensch (6) first used the term kladogenesis (or cladogenesis) to describe phylogenetic branching and anagenesis to describe the development toward higher phylogenetical levels. Certain laws of evolution were laid down that characterize the fossil record and natural populations. The law of the unspecialized states that unspecialized organisms will survive, but the overspecialized will be eliminated by changing environments (7). Furthermore, Dollo's law indicates that major evolutionary steps, once taken, are never reversed, i.e., a reptile cannot become a fish again (8). Although there are exceptions to these laws, they can be used to describe the major features of evolution.Figure 1.9. Some common evolutionary patterns as indicated by morphological changes among different lineages.
Several patterns of morphological change are also apparent during macroevolution. Divergence is the splitting of a lineage into two adaptive pathways. Chimpanzee and man are presumed to be the descendants of a common ancestor that subsequently diverged into two adaptive pathways. Radiation is a multiple divergence in which a number of lineages split from a primitive form and diverge to occupy a number of distinctive niches and regions in the biosphere. The various species of finches in the Galapagos Islands first described by Darwin were probably the result of adaptive radiation from an ancestral form that migrated from the mainland of South America (see 1.2.4, Figure 2.29 for details). Parallel evolution results when two distinct lineages undergo the same morphological changes due to adaptation to similar environments. The eye of a squid is remarkably similar to that of man although the two lineages evolved independently. This is the result of parallel evolution. And finally, convergence occurs when separate lineages become morphologically similar. The flippers of whales resemble the fins of fish. However, fish and whales are little related in the evolutionary tree. Therefore, the resemblance is due to convergence. Figure 1.9 illustrates these patterns (9). [53]
1.5.2 Synthetic Theory. The modern synthetic theory of organic evolution involves three categories: (1) the origin of life (abiogenesis), (2) microevolution, (3) macroevolution. Attempts have been made to correlate the origin of life with the physical and chemical reactions that are occurring in the inorganic world. Since the spontaneous generation of life is no longer thought to be possible under present earth conditions, evolutionists resort to the assumption that life may have occurred by the random collision of inorganic molecules on the surface of the primitive earth. These collisions were thought to be aided by physical and chemical catalysts resulting in the production of increasingly complex organic molecules by a mechanism analogous to natural selection (10). Several experiments have been set up that have simulated presumed primitive earth conditions in order to test the above hypothesis (see I.3.3, Figure 3.10; 3.11).
Much circumstantial evidence is available to support most of the contentions of the synthetic theory; however, the backbone of this huge structure is still in the process of construction. G. A. Kerkut, in the conclusion of his book The Implication of Evolution (11) summarized the current status of the synthetic theory of evolution, which he calls the "General Theory of Evolution" (macroevolution), as compared to the "Special Theory of Evolution" (microevolution) as follows:
There is a theory which states that many living animals can be observed over the course of time to undergo changes so that new species are formed. This can be called "The Special Theory of Evolution" and can be demonstrated in certain cases by experiments. On the other hand there is the theory that all the living forms in the world have arisen from a single source which itself came from an inorganic form. This theory can be called the "General Theory of Evolution" and the evidence that supports it is not sufficiently strong to allow us to consider it as anything but a working hypothesis. It is not clear whether the changes that bring about speciation are the same nature as those that brought about the development of new phyla. The answer will be found by future experimental work and not by dogmatic assertions that the General Theory of Evolution must be correct because there is nothing else that will satisfactorily take its place.Since Kerkut's statement, not much progress has been made toward the resolution of the status of the "General Theory of Evolution." Recent hypotheses attempting to account for possible genetic variabilities have placed a great deal of strain on the Neo-Darwinists who advocate the universal sufficiency of natural selection in evolution. The following section examines the evidence for the theories of evolution and attempts to evaluate the strengths and weaknesses of the theories. [54]
![]()
![]()
![]()
References 1.5
1. Goldschmidt, R. D. The material basis of evolution.
New Haven, CT: Yale Univ. Press; 1940.
2. Dobzhansky, T. Genetics and the origin of species.
1st, 2nd, 3rd ed. New York: Columbia Univ. Press; 1937, 1941, 1951.
3. Simpson, G. G. The major features of evolution.
New York: Columbia Univ. Press; 1953 (chapter 11).
4. Savage, J. Evolution. 2nd ed. New York:
Holt; 1969 (chapter 10).
5. Mayr, E. Animal species and evolution. Cambridge,
MA: Harvard Univ. Press; 1963: 481.
6. Renseh, B. Evolution above the species level.
New York: Columbia Univ. Press; 1960 (chapter 6).
7. Rensch, B. Evolution above the species level.
237.
8. Dodson, E. O.; Dodson, P. Evolution: process
and product. 2nd ed. New York: Van Nostrand; 1976: 256.
9. Dobzhansky, T.; Ayala, F. J.; Stebbins, G. L.;
Valentine, J. W. Evolution. San Francisco: Freeman; 1977: 326-27.
10. Oparin, A. I. Genesis and evolutionary development
of life. New. York: Academic; 1968.
11. Kerkut, G. A. Implications of evolution.
New York: Pergamon; 1960: 157.