2.4 Evidence from Biogeography
The study of biogeography is the categorization of the geographic distribution of fauna and flora. Careful studies of the biosphere reveal that each species of plant or animal has a certain range of existence on the earth. Jordan's rule, developed by David Starr Jordan, describes the distribution of ranges. In general, closely related species do not have identical ranges, but often their ranges are not very far apart. They are usually adjacent but sometimes separated by a barrier, such as a mountain, desert, or ocean.
Certain species of plants and animals are widely distributed throughout the world. In contrast, other species are confined to a certain location even though climatic conditions in other regions of the world seem well suited for their existence. The marsupial animals such as the opossum and kangaroo, found exclusively in North America and Australia, respectively, are examples of species with restricted distribution. In other instances, there are extreme discontinuities in the geographic distribution of a particular species or related species. The camel is found in Asia and Africa while their close relative the llama is only in South America. Other animals and plants are found in widely separated areas of the world: lungfish are known in Australia, Africa, and South America; alligators in the south-eastern [119] United States and the Yangtze River of China; and magnolias in the eastern United States, China, and Japan.
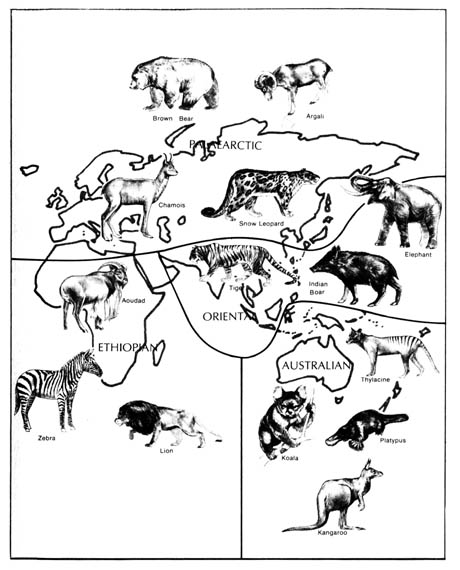
The attempts to classify the patterns of distribution of living fauna have yielded six major biogeographic realms, namely, Nearctic, Palearctic, Australian, Oriental, Ethiopian, and Neotropical regions (Figure 2.24). The realms are separated by great physical barriers. However, diverse areas within the realms have been accessible to organisms found in them throughout geological time. The regions were originally classified according to the distribution of mammals, but later it was found that the regions are valid for other kinds of fauna and flora as well.
The Nearctic and Palearctic are sometimes grouped together as the Holarctic region that includes all of Europe and Asia (north of the Himalayan and Nan Ling mountain ranges), Africa (north of the Sahara desert), and North America (north of the Mexican Plateau). The animals found in the Nearctic (North America) and Palearctic (Old World) exhibit differences at the species and generic levels only. Representative animals found in the Nearctic realm are mountain goats, prairie dogs, opossums, skunks, raccoons, blue jays, and turkey buzzards. Some of the indigenous animals found in the Palearctic realm include goats, moles, deer, oxen, sheep, robins, and magpies.
The Australian region includes Australia and the associated islands. It is characterized by a predominantly marsupial fauna and the complete absence of any native placental mammals, other than bats and some rodents.
The Oriental region is separated from the Australian region by an imaginary dividing line known as Wallace's Line. It separates the islands of Bali and Lombok that are only 20 miles (32 km.) apart. It also goes through the Macassar Straits and passes to the east of the Philippines. The mammals found on both sides of this imaginary line are drastically different. The orangutan, black panther, tiger, water buffalo, Indian elephant, gibbon, and tarsier are characteristic animals of the Oriental region. However, the kangaroo, koala, and platypus are characteristic animals of the Australian region.
The part of Africa south of the Sahara and the island of Madagascar constitute the Ethiopian realm. Animals found exclusively in this region are the giraffe, aardvark, chimpanzee, gorilla, zebra, and hippopotamus.
Finally, the Neotropical region consists of South and Central America,
southern Mexico, and the West Indian islands. The following animal's make
up the distinctive fauna in this realm: the alpaca, tinamous puff bird,
capybara, bloodsucking bat, sloth, llama, prehensile-tailed monkey, tapir,
and anteater.
The distinctive habitats (ecological zones) within (e.g., lakes) and between [120] (e.g., oceans) the biotic regions are barriers that affect the geographic distribution of fauna and flora. Because of the climatic difference in the attitudinal and latitudinal zones, plants are distributed according to their abilities to adapt to each habitat (Figure 2.25). Thus, ptarimigans and the varying hare are found in the higher mountains of the western United States as well as in the arctic and subarctic lowlands of Canada and [121] Alaska because the habitats are similar in each location. In Europe, the mountain hare is found in the mountains ranging from the east (Caucasus and Ural mountains) to the west coast and also in the arctic lowlands but not in the intervening lowlands. [122]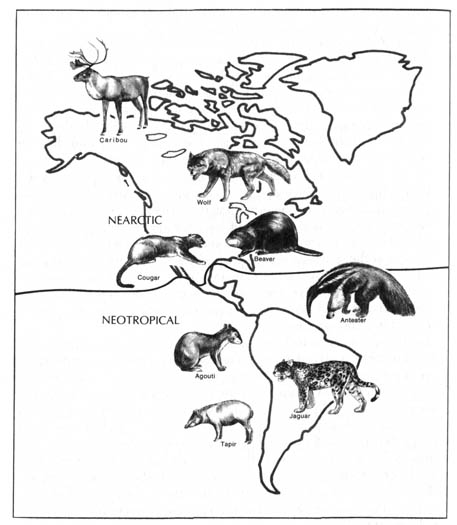
Figure 2.24. The biogeographic realms of the world. Reprinted, with permission, from Villee, C. A. Biology. 7th ed. Philadelphia. W. B. Saunders Co.; 1977. © 1977 by the W. B. Saunders Co.
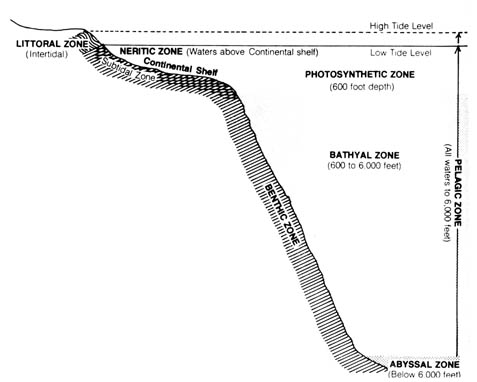
Similarly, the different ecological zones (habitats) of the ocean each possess its own distinct flora and fauna (Figure 2.26). The narrow strand [123] on each shoreline that is alternatively covered and exposed by the tide is the littoral (intertidal) zone. The continental shelf usually slopes beyond this zone, forming continental islands at its higher portions. The shallow seas that cover the continental shelves, less than 600 feet deep, are the neritic zone. When the ocean floor drops off abruptly at the outskirt of the continental shelf, several zones of great depths are created. The surface layer of seas to a depth of as much as 600 feet is the pelagic zone where water is well aerated, lighted, and subject to wave action. The deeper seas, to a depth of 6000 feet, comprise the bathyal zone. The lowest part of the sea, the abyssal zone, is extremely cold, quiet, devoid of sunlight, and can sustain only a few profoundly modified and adapted organisms. This is in marked contrast to the littoral zone, which is the most richly inhabited area in the ocean. The deep sea basins form barriers that affect the geographical distribution of the marine fauna and flora found within the various continental shelves.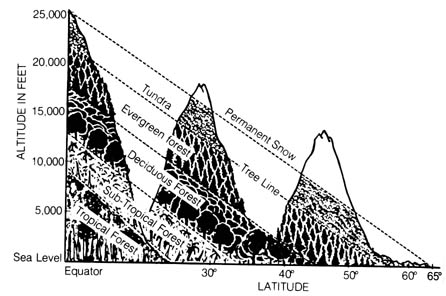
Figure 2.25. Comparison of latitudinal and attitudinal life zones of plants in North America. Reprinted, with permission, from Moore, J. N.; Slusher, H. S. editors. Biology. Grand Rapids, MI: Zondervan Publishing House; 1970.Figure 2.26. Ecological zones in the ocean. Reprinted, with permission, from Moore, J. N.; Slusher, H. S. editors. Biology. Grand Rapids, MI: Zondervan Publishing House; 1970.
The distribution of animals in the six biogeographic areas has been an intriguing phenomenon for biogeographers ever since the inception of this branch of science. Many theories have been postulated to account for this phenomenon based on a uniform geological outlook with minor modification throughout the earth's history (1). However, these theories have not been very convincing. In exploring the migration theory, it is difficult to understand how the migration of reptiles and amphibians could have led to the deposition of the same amphibians and reptile fossils that are found on Antarctica, South America, South Africa, and the Indian continents when these areas are separated by the barriers of oceans and mountain ranges.
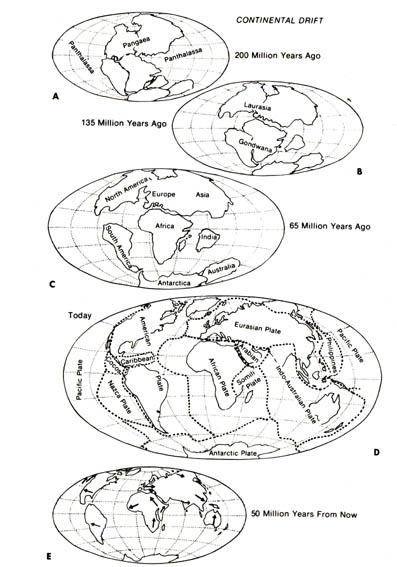
The continental drift theory has been put forth to account for some
of the geographic distribution problems. This theory, which dates back
to 1910 (10), states that from the Paleozoic era until late in the Mesozoic
era only two major land masses existed, and the two, Gondwanda and Laurasia,
were in contact. Gondwanda centered around the South Pole while Laurasia
extended well into the northern hemisphere. During the Cretaceous era,
Gondwanda gave rise to the southern continents by drifting away: Subsequently,
the southern continents split to form Africa, South America, Australia,
and Antarctica, as well as the Arabian and Indian peninsulas and the major
Pacific islands such as New Zealand and Madagascar. Laurasia, on the other
hand, broke up and formed North America, Europe, and Asia (Figure 2.27).
The continental drift hypothesis was unattractive when it was first proposed because no known mechanism could account for the drift. However, during the 1960s, the discovery of terrestrial paleomagnetism [125] sparked new interest in the continental-drift theory. Terrestrial paleomagnetism holds that the continents of the world have changed their positions relative to the earth's magnetic pole during its history (2). The plotting of Paleozoic paleomagnetic pole positions indicates that the southern continents and India were clustered together in the far south (9).Figure 2.27. Continental drift. A, The supercontinent Pangaea of the Triassic period, about 200 million years ago. B, Breakup of Pangaea into Laurasia (Northern Hemisphere) and Gondwana (Southern Hemisphere) 135 million years ago in the Cretaceous period. C, Further separation of land masses, which occurred in the Tertiary period, 65 million years ago. Note that Europe and North America are still joined and that India is a separate land mass. D, The continents today. E, Projected positions of the continents in 50 million years. Reprinted, with permission, from Villee, C. A. Biology. 7th ed. Philadelphia: W. B. Saunders Co.; 1977. © 1977 by the W. B. Saunders Co.
A mechanism based on circumstantial evidence, namely, plate tectonics, has been postulated to explain continental drift (3). The theory essentially states that the earth's crust is composed of a rigid outer shella lithosphere, resting on an underlying and less rigid asthenosphere. The outer shell is broken up into some 6 (4) or 10 (5) major plates. These plates and subplates of varying shapes and sizes are in a state of flux relative to each other, presumably as a result of the forces and processes occurring in the asthenosphere. The mechanisms by which plates move relative to each other is not clear. It is conceivable that they are being pushed, pulled, driven by gravitational forces, or carried by convection units into the mantle of the earth. The boundaries of the major plates coincide with the regions of major tectonic activities (e.g., San Andreas fault) of the lithosphere, such as earthquakes.
With the increasing popularity of the continental-drift theory, the
biogeography of the world could be reinterpreted. Before the advent of
evolutionary thinking, it was thought that similar fauna and flora in isolated
areas of the world represented independent and unique creations. However,
it was intriguing that animals and plants were in exclusively one region
while equally suitable habitats in other areas were devoid of any trace
of the same organism. No one was able to offer a satisfactory reason for
the distribution patterns. However, a study of the distribution of bryophytes
(mosses and liverworts) seemed to fit nicely into the scheme of continental
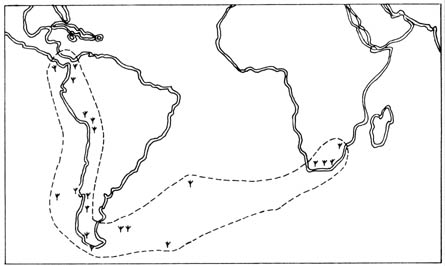
drift (6). Clasmatocolea vermicularis, a species
of leafy liverwort, was found to be distributed along the east coast of
South Africa and the west coast of South America, as well as Central America
and some islands (Figure 2.28). Since the plant is known to have a limited
capacity of dispersal, it was next to impossible for it to cross the Atlantic
Ocean in order to populate the two continents. Therefore, the distribution
was most likely the result of scattering of the species at the south coast
of Gondwanda when it split into the present continents.
Figure 2.28. The distribution of Clasmatocolea vermicularis (
) in South Africa, South and Central America as well as some islands.
In summary, the concept of natural selection, which gained popularity
with the publication of Origin of the Species by Darwin, provides
a good explanation of the diversification of species into the different
habitats of the biosphere. Darwin's concept of natural selection emerged
from his observations that began in 1831 when he became a naturalist on
the H. M. S. Beagle that sailed from England to South America as
part of a [126] survey of continental coastlines. Darwin visited the various
isolated islands off the coast of Ecuador, and on these oceanic islands,
he found a large number of endemic species of finches that are closely
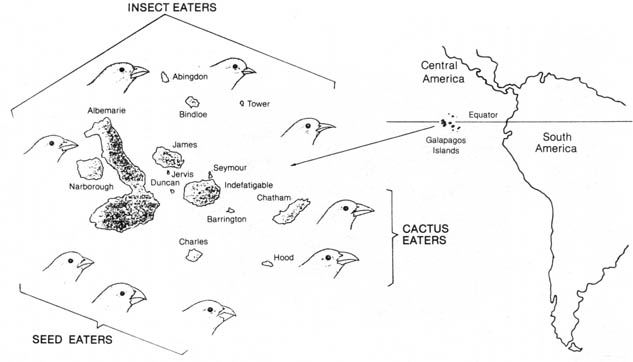
related with the finches on the South American mainland (Figure 2.29).
Certain features of the birds are distinct and can be related to the habitats
of the various populations. The most striking examples were the sizes and
shapes of the beaks of endemic finches that seemed to be correlated with
their diets. Darwin suggested that all these varieties of finches were
descendants of an ancestral species that immigrated to the islands from
the mainland. After the offspring of the ancestral species became numerous,
they outstripped the food supply. By a process of natural selection, the
variant individuals with slightly better-equipped beaks were able to survive
in distinct parts of the islands, according to the type of food available.
Over the course of time variant forms of finches came to occupy distinct
niches in the environment (7, 8).
It was also intriguing for Darwin to observe that over half of the more
than 300 plants in the Galapagos Islands were endemic. Yet, all of these
plants show close relationship to South American plants. However, the mainland
and the islands have totally different climatic and geological characteristics.
Therefore, he postulated also that the varieties of plants on the islands
descended from the introduction of the ancestral plants from the mainland,
and the subsequent changes came by natural selection (7).
[127]
Figure 2.29. The location of the Galapagos Islands and their distribution of birds according to bill structures. Reprinted, by permission, from Dodson, E. 0.; Dodson, P. 2nd ed. Evolution, process and product. New York: Van Nostrand Co.; 1976. © 1976 by Litton Educational Publishing, Inc.
[128] On this basis, Darwin suggested that the biogeography of organisms
can be explained by assuming each group of organisms after originating
in the major regions of the world, spread to occupy as much space as it
could, to be stopped only by the natural selective factors of physical
and climatic barriers and competition with other organisms. Thus, he explained
the similarities of the plants in different habitats by the assumption
that they were from the same origin and underwent morphological changes
after migration.
Theories of biogeography incorporate Darwin's assumption and further postulate that each species of fauna and flora originated only once at the "center of origin," and from there it diversified into different areas of the same region by natural selection. Thus, when the continents split apart, the species diversified separately by natural selection as the land drifted.
Based on the current assumptions of biogeographers, the occurrence of similar organisms in different biogeographical realms does not have to be attributed to separate creative acts of God. This could have resulted by continental drift, following the appearance of a species at the common "center of origin" through the creative act of God.
![]()
References 2.4
1. Dodson, E. O.; Dodson, P. Evolution, process and product. New York: Van Nostrand; 1976: 403-13.