3.3 Evaluation of Macroevolution (continued)
3.3.2 Rational Incoherency
a) Insufficiency of Natural Selection to Account for Macroevolution. Evolution above the species level has not been satisfactorily accounted for by the mechanism of natural selection even though it does explain nicely the phenomena of microevolution. This quote is from a recent text on evolution (18):
The process of natural selection, acting upon the sources of genetic variability that reside in the gene pools of species, is clearly adequate to produce, preserve, and accumulate the sorts of changes that lead one species to another. There is a voluminous body of theory and evidence to explain the origin of species through microevolution.Various concepts have been proposed to account for macroevolution. The following sections will review the concepts of neutral mutation, regulatory mutation, systematic mutation, and species selection.
The differences between distantly allied species are profound, however .... The differences between such species are in fact so impressive that some investigators have suggested that they have arisen through mechanisms distinct from the microevolutionary processes of adaptation. This has led to some of the major controversies in evolutionary theory.
(1) Concept of Neutral Mutation. As mentioned earlier in 1.1.4, the classical mutation theory was popular in the earlier part of the twentieth century until the elaborate work of Dobzhansky and others who presented a strong case for evolution by natural selection. The two theories predict different degrees of genetic variability in natural populations. [216] The classical theory predicts that most individuals in the natural population are homozygous since natural selection serves to eliminate all but a few mutations that are in the form of heterozygotes. Therefore, genetic variability in natural populations, according to the classicists, is minimal.
The Neo-Darwinist, or the selectionist, on the other hand, predicts exactly the opposite results. Since natural selection works on gene mutations to cause changes in gene frequencies, many individuals in natural populations, which are presumably undergoing evolutionary change, would be heterozygotes that arise from mutations but are selected by overdominance (see I.3.2.1). According to selectionists then, genetic variability in natural populations is the rule instead of the exception.
Before the evidence from the studies of molecular evolution was available, only mutations that had drastic phenotypic effects were analyzed, and the phenomenon of genetic variability in natural populations could not be empirically detected. Therefore, although examples of polymorphism (i.e., situations where the members of a natural population can be sharply categorized into two or more relatively common phenotypes that are determined by commonly occurring alleles at a particular gene locus, such as blood types) were recognized in genetics early in the science, it was impossible to tell whether these represented special cases or a widespread phenomenon.
In the late 1960s, workers began to employ the technology of molecular biology to tackle the problem of genetic variability in natural populations. By the application of electrophoretic techniques, it was found that enzyme and protein polymorphism both in Drosophila (19, 20) and the human (21) is a common phenomenon. Similar studies have been carried out in plants (22, 23) and in animals, including protozoans, mollusks, arthropods, bryozoans, echinoderms, and vertebrates (24). It has become clear that this phenomenon is ubiquitous at least in the natural populations of animals. The discovery of the widespread polymorphisms together with the apparent constant rate of molecular evolution (see I.3.3.1.d) gave the classicists new impetus, and they postulated a new concept of neutral mutation in reformulating their theory. Presently, they are called the neo-classicists or panneutralists as contrasted with the Neo-Darwinian school of selectionists.
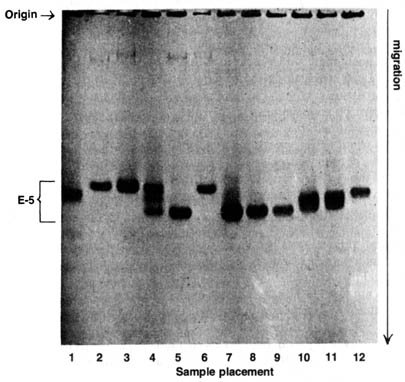
There are two parameters in the description of enzyme or protein polymorphisms in natural populations: (1) percentage of loci (alleles) that are polymorphic, and (2) percentage of loci (alleles) that are heterozygous. Each parameter can be accurately estimated by analyzing the gel electrophoretic pattern (Figure 3.19) showing a typical analysis of enzyme [217] polymorphism. The electrophoretic pattern can allow the differentiation of polymorphic alleles and can show if an organism is homozygous or heterozygous at a genetic locus.
The phenomenon of polymorphisms and heterzygosity is illustrated in
a study of enzymatic differences within a single species. Esterase-5
is
an enzyme known to be synthesized by the gene locus 5 in a particular chromosome
of Drosophila pseudoobscura. Sample 1 is a homozygous standard strain,
as indicated by a discrete protein band synthesized by two identical alleles
on locus est-51.00. The superscript stands for the relative
electrophoretic mobility of the protein specified by the allele. Samples
2, 3, and 6 have a slower migrating protein, while sample 5 moves faster
than the standard. Sample 4 shows three bands, two identical with the fast-
and slow-moving bands and one intermediate between [218] them. Samples
7-12 contain other proteins that are irrelevant to the present discussion.
By looking only at the mobilites and the discreteness of the protein bands
in sample 5 and samples 2, 3, and 6, one can infer that the two homozygotes
est-51.12
and est-50.95 are represented by the fast- and slow-moving
bands respectively. Thus, we can conclude that the esterase-5 locus
is polymorphic, because different homozygotes can be detected according
to their electrophoretic mobilities from the natural population of D.
pseudoobscura.
The three bands shown in sample 4 can be identified as a heterozygote est-50.95 / est-51.12 . The presence of the intermediate band in sample 4 indicates that esterase-5 is a dimeric enzyme (consisting of two polypeptides). Therefore, the heterozygote, est-50.95 / est-51.12 can produce three dimers: (1) homodimer 0.95-0.95, with the same mobility as the dimer made by the homozygote est-5^0.95; (2) homodimer 1.12-1.12, with the same mobility as the dimer made by the homozygote est-51.12 / est-51.12; and (3) a hybrid dimer 1.12-0.95, with a mobility halfway between. This heterozygosity of sample 4 is confirmed by the genetic analysis of individuals from the strain with extracted protein of sample 4 that segregates into two different alleles. Thus the electrophoretic pattern easily reveals the heterozygosity of a locus regardless of the dominance or recessiveness of the alleles because the product of each allele is examined.Figure 3.19. Adult esterases from Drosophila pseudoobscura. Sample 1, standard strain est-51.00/est-51.00; samples 2 and 3, est-50.95/est-50.95; sample 4, est-50.95/est-51.12; sample 5, est-51.12/est-51.12; sample 6, est-50.95/est-50.95. Reprinted, with permission, from Lewontin, R. C. The genetic basis of evolutionary change. New York: Columbia University Press; 1974.
Even when an enzyme is monomeric (consisting of only one polypeptide), the heterozygote is easily detected also for it makes two different forms of the enzyme. Each form corresponds to one of the homozygotes in mobility despite the absence of the "hybrid" molecule as seen in sample 4. The different enzyme forms produced by different alleles at the same locus are called allozymes. The difference of mobility in the various protein bands represents only one-third or one-half of the amino acid changes in the polypeptides that are responsible for the polymorphism. Many amino acid substitutions involve no change in net charge of the protein and are not detected by the change in electrophoretic mobilities. Therefore estimates of polymorphisms and heterozygosities are only for the "lower limits" of what the case actually is. From these data it is estimated that in sexually reproducing species of animals, one-third of their genes are polymorphic, and 10% of the loci of the individuals within the species are heterozygous (25).
The principles set forth by the neo-classicists to account for molecular evolution are fivefold (26): (1) For each protein, the rate of evolution in terms of amino acid substitutions is approximately constant per site per year, as long as the function and tertiary structure of the protein [219] molecules remain essentially unaltered. (2) Functionally less important molecules or parts of molecules evolve (in terms of mutant substitutions) faster than more important ones. (3) Those mutant substitutions that cause less disruption in the existing structure and function of a molecule (conservative substitutions) occur more frequently in evolution than do more disruptive ones. (4) Gene duplication must always precede the emergence of a gene having a new function. (5) Selective elimination of definitely harmful mutants and random fixation of selectively neutral or very slightly harmful mutants occur far more frequently in evolution than the positive Darwinian selection of definitely advantageous mutants.
The first condition is somewhat substantiated by the development of
the molecular clock (see I.3.3.1.d, Figure 3.16) and the apparent
constant rate of evolutions in some proteins (see Figure 3.14).
Despite some disparities in the rate of evolution in some proteins, the
rate of neutral mutations is largely unknown because of limited data. However,
it can be calculated that the rate of gene substitution for neutral genes
is equal to the mutation rate, irrespective of population size (27,13).
Therefore, if mutation rate remains constant, the rate of evolutionary
change for a given protein would also occur with constant probability.
The second condition seems to be well documented by the fact that fibrinopeptide
and proinsulinpolypeptide C, both relatively useless proteins, evolve 18
and 11 times respectively, faster than cytochrome c, which is an
essential protein in the energy transport mechanisms (28). The evolution
of hemoglobin ![]() and
and ![]() chain lends support to the third condition. The surface part of the molecules
evolves nearly 10 times as fast as the functionally important heme pocket.
chain lends support to the third condition. The surface part of the molecules
evolves nearly 10 times as fast as the functionally important heme pocket.
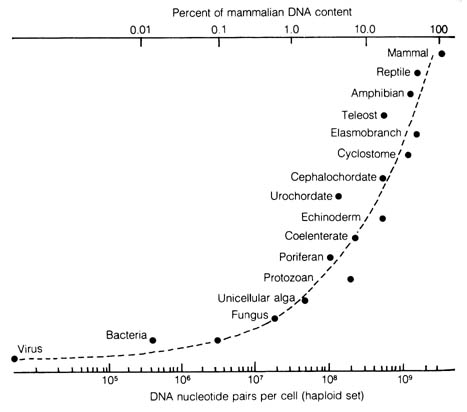
The fourth condition of gene duplication is a mechanism proposed initially to explain protein evolution of hemoglobins and myoglobins that share many common amino acid sequences and similar functions. It seems to be the most appealing mechanism to account for the evolutionary acquisition of a large amount of DNA in higher organisms (Figure 3.20). However, the theory of gene duplication has no empirical documentation. The lack of apparent selective values of most of the polymorphisms observed in natural populations are taken to be indicative of the failure of natural selection to maintain genetic variability of these loci. Last, natural selection is taken by panneutralists as the editor, rather than the composer, of the genetic message as claimed by the selectionists.
The controversies between panneutralists and Neo-Darwinian selectionists
can be represented in their disagreements over the interpretation of two
phenomenathe apparent constant rate of protein evolution and the widespread
protein polymorphism. First, the panneutralists claim [220] that the apparent
constant rate of protein evolution (which may be subject to different interpretations)
is strong evidence to support the hypothesis that it is the result of neutral
mutation, the rate of which determines the rate of amino acid substitution
by random genetic drift. However, the selectionists have to make very specific
assumptions on mutation rates, selective advantages, and effective population
sizes in order to explain the apparent constancy of protein evolution.
On the one hand, selectionists question the constancy of protein evolution.
On the other hand, they stress that amino acids in a protein sequence in
a species have been selected in the course of evolution because they are
best adapted to meet the particular features of external environment. Selection
of an amino acid [221] sequence of a protein also depends on the activities
of the other genes present.
In considering the second phenomenon, panneutralists maintain that the widespread polymorphism in natural populations is the result of mutations that are neither beneficial nor harmful and are fixed by random genetic drift. Natural selection is mainly responsible for the elimination of harmful mutations and only occasionally establishes rare advantageous mutations. Selectionists argue that the heterozygotes are responsible for carrying the mutations and are selectively more advantageous (hybrid vigor or overdominance) than the homozygotes. Therefore, protein polymorphisms are the expressions of a stable genetic equilibrium maintained by balancing selection (see I.3.2.1.c).Figure 3.20. The minimal amount of DNA that has been observed for various species in the types of organisms listed. Each point represents the measured DNA content per cell for a haploid set of chromosomes. The ordinate scale and the shape of the curve is arbitrary. Reprinted, with permission, from Nei, M. Molecular population genetics and evolution, New York: Elsevier & N. Holland; 1975.
Both theories seem to account for some of the observations of molecular evolution, but both have serious shortcomings. The panneutralists have to document their assertion of the constant rate of protein evolution more vigorously in light of the disparities observed in some known cases. In addition, the proportions of allozyme polymorphisms showing selective values are considerable and cannot be easily ignored.
The balancing selection theory is plagued with the conflict between "hybrid vigor" and "inbreeding depression." If a large percentage of natural populations are heterozygous as maintained by balancing selection, a significant percentage of homozygotes would be derived by the inevitable inbreeding of heterozygotes, and the genetic variability of the populations would be reduced accordingly. However, this was not observed. Moreover, heterozygotes produced by bisexual organisms seem not to be necessary for the maintenance of protein polymorphisms for self-fertilizing plants (22) and bacteria (29). Parthenogenic species of animals (24) are also shown to be polymorphic in many loci.
These data place severe strains on the selection theory. The rarity of observable hybrid vigor, except the well-studied sickle cell anemia (see 1.2.6.2; 3.2.1.c), also does not aid the selectionist's position. At the present time the controversy between the selectionists and panneutralists goes on with no solution yet in sight. It is fair to conclude that the foundation of macroevolution based on natural selection is seriously shaken by the panneutralists.
(2) Concept of Regulatory Mutation. The concept that major anatomical changes are the result of mutations affecting gene expression (regulatory mutations) was proposed to account for the apparent inconsistency between molecular and organismal evolution (30) (see I.3.2.1.d). According to this hypothesis, small differences in the time of activation or in the level of activity of a single regulatory gene could in principle [222] influence considerably the systems controlling embryonic development. The organismal differences between chimpanzee and humans would then probably result chiefly from genetic changes in a few regulatory systems that are hardly detectable due to the difficulties involved in the purification and identification of regulatory proteins.
Regulatory mutations can occur in two ways. First, nucleotide substitution can affect a regulator gene that affects production but not the amino acid sequence of proteins in that operon (see I.2.6, Figure 2.52). Second, chromosomal rearrangements by inversion, translocation, duplication, deletion, or transposition may be responsible for the change in genetic expression without damaging the amino acid sequence of the gene products; however, the biochemical mechanisms behind these changes are obscure.
The regulatory mutation concept is purely speculative, for there is no empirical evidence to support it. The operon model is well documented in the bacterial system, but it has not been unequivocally identified in the eucaryotic genome due to the following four reasons:
1. DNA in the eucaryotic chromosome is wrapped in chromosomal proteins that may play important roles in the regulation of gene expression. The presence of these protein makes the identification of genes involved in the operon very difficult.Realizing the complexity of the eucaryotic genome (31), scientists have yet to work out the regulation of gene expression. Therefore, the concept of regulatory mutation can be treated only as a speculation that is difficult to test empirically. At the present time it is very much doubted whether the effects of regulatory gene mutation observed in bacteria (32) can be applied to the eucaryotic system.2. Genomes of higher organisms contain various classes of highly repeated DNA with virtually unknown functions.
3. A large part of the highly repeated DNA is apparently nonfunctional because it does not transcribe any RNA.
4. The nucleus of an eucaryotic cell contains heterogeneous nuclear RNA with high complexity (10 times that of cytoplasmic mRNA) that later becomes mRNA in the cytoplasm. Several other low molecular weight species of RNA with unknown function are also found in the nuclei.
(3) Concept of Systemic Mutation. The late Richard B. Goldschmidt (1878-1958), geneticist at the University of California, has expressed frustration in trying to account for the macroevolutionary development of many structures in higher organisms on the bases of the mechanisms of microevolution alone. He believed that the Neo-Darwinian mechanism (the accumulation of micromutations under the [223] influence of natural selection) was largely restricted to subspecific differentiation within species and that the decisive step in the formation of new species involves an entirely different genetic process called systemic mutation (33, 34).
Goldschmidt's reasoning on systemic mutation is threefold. First, if microevolution gives rise to new species according to different stages of geographic isolation, it should be possible to observe an entire series of geographically isolated subspecies with the terminal one representing the beginning of a new species. Goldschmidt expected to find these series of geographically isolated subspecies of closely related species blending into one another, but he cited many examples in which this blending did not occur. He treated a species as an interbreeding or potentially interbreeding population, and he claimed that many controversial cases of speciation depend in part on the purely morphological definitions of a species that do not take genetic aspects into account. Therefore he believed that good species are always separated from their nearest relatives by a bridgeless gap.
Goldschmidt's second point is that natural selection acting via geographical isolation is believed to cause the accumulation of enough genetic difference in the isolated subspecies so that it eventually becomes a new species and is distinct from the parent species. But Goldschmidt documented in many instances that long isolation did not produce more than subspecific variations. He cited a race of the gypsy moth Lymantria dispar that has been isolated on the island of Hokkaido (North Japan) since the early Tertiary period, yet in the intervening 60 million years only subspecific differentiation has occurred. He also pointed out that seasonal varieties within a race of the butterfly Papilio may be greater than the variation between races of Papilio butterflies at any one time.
Goldschmidt's third point is that the rate of evolution directed by natural selection is too slow and subtle to account for the existing varieties of plants and animals. Neo-Darwinian theory demands that only very minor mutants subject to very slight selection pressure are significant in evolution. If one defines selection pressure as the loss of survival value, then in a population with 1000 individuals of AA genotype and 999 individuals of aa genotype who reproduce, the selective pressure against a is 0.001. According to mathematics set up by J. B. S. Haldane (35), if a selection pressure of this magnitude is operating "in favor" of a new gene that arose by micromutation present in frequency of one in a million, it will take almost 12,000 generations to increase the gene frequency to 2 in a million if the favored gene is dominant, and 322,000 generations if the favored gene is recessive. Goldschmidt inferred also from the lack of [224] genetic differentiation of natural populations above the species level such as in the case of Lymantria, which was under prolonged and complete isolation, that natural selection is ineffective in speciation.
Goldschmidt therefore advocated a wholesale chromosomal rearrangement that he called "the systemic mutation" as the novel genetic process to account for speciation. Such a drastic chromosomal rearrangement is supported by observations of Drosophila chromosomes. While natural selection usually eliminates such individuals who arise from systemic mutation, occasionally it may allow them to propagate as "hopeful monsters" under special circumstances.
The concept of systemic mutation lacks sufficient empirical documentation. The only observable example seems to be polyploidy in plant speciation, and this cannot be generalized to represent all living organisms. However, the body of highly pertinent evidence amassed by Goldschmidt to support his contention that the mechanisms of microevolution fail to account for macroevolution has prompted a reevaluation of the roles played by natural selection in the process of evolution.
Systemic mutation has also found its resurgence in a new theory called punctuated equilibrium. This theory is advocated by Stephen Jay Gould of Harvard University and Niles Eldredge of the American Museum of Natural History of New York (36). The essence of this theory is that during evolutionary periods individual species remain virtually unchanged. Speciation occurs only as "punctuations" caused by abrupt events at which a descendant species arises from the original stock. This view finds increasing acceptance among paleontologists who are dissatisfied with the imperfection of the fossil record, which lacks many transitional forms.
The abrupt appearance of many animal body plans ranked as phyla and classes approximately 500 million years ago has received more attention in light of newly discovered fossil beds. It has drawn renewed interest in what is so called the Cambrian explosion since the 1980s. The top down pattern in which higher levels of biological hierarchy appear first, or Disparity precedes Diversity, contradicts the Darwinian prediction that lower forms should emerge before higher ones.(43) Thus punctuated equilibrium has become a more dominant biological theory. However, it still lacks the experimental status of a testable mechanistic explanation.
(4) Concept of Species Selection. Reacting to arguments of opponents of macroevolution, modern evolutionists have tried to reiterate their conviction that the process of natural selection is responsible for both microevolution and macroevolution. Yet the failure of natural selection (acting on individuals within a population) to account for the major features of macroevolution as represented by the fossil record has led to the formulation of the concept of species selection (37).
It states that the random process of speciation favors species that speciate at high rates and survive for long periods; therefore, they tend to leave many daughter species. The concept was based on several extrapolations from the fossil record: (1) The time a new species appeared to its extinction or its pseudoextinction by gradually evolving into another species is between six to seven million years. (2) The duration time for mammalian species are shorter than those of some marine vertebrates with the major orders of mammals arising from their primitive ancestors within a span not exceeding [225] 12 million years. (3) During an interval not greatly exceeding five million years, a new aquatic-animal family Limnocardiidae arose and developed over 30 new genera representing five subfamilies with members showing great morphological diversity. (4) The presence of "living fossils" such as the linguloid, brachipods, monoplacophoran mollusks, rhynchocephalian reptiles, mytilid and pinnid bivalve mollusks, sclerosponges, and the lungfishes indicate little or no evolutionary changes over a period of hundreds of millions of years after their origination. These phenomena are not adequately explained by the gradualistic accumulation of micromutations selected by geographic isolation (allopatric speciation), but instead they demand a rapid evolutionary mechanism to account for an initial fast apparent evolution rate followed by little if any change.
The concept of species selection bears a certain resemblance to the concept of sympatric speciation (see I.1.5) in that a sudden rapid mechanism is responsible for the formation of new species. It is analogous also to the process of natural selection in that species are selected on the basis of their ability to resist extinction and to form a new species. In contrast, natural selection occurs if individuals within a population exhibit genetic variabilities caused by mutation and recombination. These individuals are selected according to their abilities to survive and according to their rates of reproduction. While the concept of species selection may account for the apparent rapid evolution of certain lineages represented in the fossil record, it has no empirical documentation such as that of the mechanism of natural selection in microevolution. Once again the mechanism of natural selection fails to account for the major features of macroevolution in the concept of species selection.
Recently, with the discovery of the uniqueness of Archaebacteria in rRNA sequence and by comparative studies with well-characterized molecular systems, cell walls, lipid compositions and features of the transcriptional and translational machineries, the three domains of life, namely Archaea, Bacteria and Eukarya, has become the currently accepted paradigm in the field of molecular taxonomy.The three domains arise from an ancestral cellular community (progenotes) undergoing high rates of mutation and lateral gene transfer. The three domains crystallize into separate cellular communities in this situation, becoming distinctive and refractory to lateral gene transfer. In this fashion the domains are monophyletic and entirely independent. The implication of polyphyletic origins of the three domains of life from a universal pool of progenotes seems to demand a mechanism beyond the realm in which Darwinian natural selection canoperate.(44, 45, 46, 47, 48)
All in all, the idea of Darwinian evolution is still venerated as the most comprehensive theory in biology. However, the concept of natural selection, by which the theory was given a scientific basis, is being gradually abandoned by the more radical biologists as the major mechanism that can account for the features of macroevolution. In addition, a recent conference on macroevolution has epitomized the growing contention that macroevolution occurs by a mechanism other than natural selection (36). The Darwinian evolutionists are no longer the dominating voice in the scientific debate on evolution. R. C. Lewontin has summed up his evaluation of natural selection as follows (38):
During the last few years there has been a flowering of interest in evolution by purely random processes in which natural selection plays no role at all. If the empirical fact should be that most of the genetic change in species formation is indeed of this "non-Darwinian" sort, then where is the revolution that Darwin [226] made? The answer is that the essential nature of Darwinian revolution was neither the introduction of evolutionism as a worldview (since historically that is not the case) nor the emphasis on natural selection as the main force in evolution (since empirically that may not be the case), but rather the replacement of a metaphysical view of variation among organisms by a materialistic view.In other words the only contribution that Darwin's natural selection theory made is in the form of an empirically testable mechanism that causes the diversification of genetic variability as exemplified by the process of microevolution. Macroevolution, then, is a very speculative theory that is becoming gradually divorced from the well-documented concept of natural selection (42).
b) Chance as the Teleological Explanation of Evolution. Aristotle has categorized four levels of explanation of an event: material cause, efficient cause, formal cause, and final cause. A sculpture can be used as an example. The material explanation of it is the stone or wood from which the sculpture is made, i.e., the sculpture is a piece of stone or wood. The efficient cause is the force, the act of carving, that forms the figure of the sculpture, i.e., the sculpture is a carved stone or wood. The formal cause is the pattern after which the sculpture is carved, i.e., a sculpture is a carved stone statue of man. The final cause is the purpose of the existence of the sculpture, i.e., the sculpture is a carved stone statue of Abraham Lincoln, commemorating his work as president of the United States.
According to Webster's Third New International Dictionary, the use of design, purpose, or utility as an explanation of any natural phenomenon is known as teleology. Thus, a teleological explanation of an event is equivalent to Aristotle's final cause. The English theologian William Paley argued eloquently in his Natural Theology that the intriguing features of nature evidence the design of the Creator. He cited the example of the human eye. Paley points to the fitting together efficiently and cooperatively of the lens, retina, and brain; enabling humans to have vision; as conclusive evidence of the design of an all-wise Creator. Thus the functional design of organisms and their features are taken as evidence of the existence of the designer.
Darwin rejected the notion of a designer and argued that the directive organization of living things is the result of a natural processnatural selection. In other words, he stressed only the material and efficient causes of the features of the organisms and tried to bring the origin and adaptation of organisms into the realm of empirical science. Evolutionists maintain the natural process can be accounted for by physiochemical [227] parameters; therefore, there is no need to resort to the design of a Creator or external agent. Nevertheless, they invoked natural selection as an agent capable of providing a purpose for the existence of certain features of organisms. Evolutionists have on the one hand regarded the teleological explanation of natural phenomena as untestable scientifically and thus untenable; but they have asserted, on the other hand, that chance, directed by natural selection, is the ultimate explanation of the necessity of evolution (39).
The results of natural selection as exemplified by the adaptive feature of the hand of humans, the wings of birds, and other biological structures or behaviors have been treated as the reasons why they exist at all. To put it more precisely, according to the evolutionists, the reason why there are streptomycin-resistant mutants in a population of E. coli bacteria is that they can propagate in the presence of the drug, whereas streptomycin sensitive bacteria cannot. Since streptomycin-resistant mutation arose spontaneously, the selection by the drug simply facilitates the differential multiplication of the mutant at the expense of the nonmutated bacteria. Therefore, according to the evolutionists, the "music of the biosphere" is composed of the unaided "noise of natural selection" feeding on "chance" alone (39).
Chance mutations can be subjected to the following fates: (1) selection by favorable natural environments thus providing the raw materials for further evolution, and (2) fixation by random drift or elimination by adverse conditions thus moving to oblivion. The assertion that the present biota is "entirely" the result of successful evolution of chance events by the process of natural selection is purely a posteriori, since natural selection is also known to cause extinction. The same assertion can be made to describe a hypothetical barren earth as the result of natural selection of unsuccessful chance events. Therefore evolutionists, while stressing the material and efficient causes of evolution in the mechanism of natural selection, have yet to come up with a valid counterargument to explain why chance alone can be in such marvelous harmony to produce the orderly array in the biosphere. Could not chance equally cause disruption of the whole structure? Both of these phenomena would be equally probable conditions implicit in the use of the term "chance."
c) Empirical Unfalsifiability of the Theory of Organic Evolution. After the triumphant Centennial Celebration of Darwinism in 1959, a mathematical and philosophical debate continued into the 1960s regarding the logical coherency of Darwin's concept of natural selection. The arguments focused on the circular reasoning of Darwin's premise of the survival of the fittest. Darwin did not provide any objective criteria to [228] identify the fittest other than observing the survivor (40).
Evolutionists tried to get around the apparent tautology of their theory by redefining natural selection to mean differential reproduction that gives rise to changes in gene frequencies. In other words, a particular genetic variant confers higher fitness in a particular situation. Therefore it will leave more offspring in the course of time. The catch, however, is still present; i.e., the conclusion is essentially part of the premise, since the fitness is measured by the capacity to leave more offspring. Grease summarized this change as follows: "What have we? One more tautology: well, after all what survives survives . . . . When the theory [of natural selection] is summed up in a formula for measuring differential gene ratios, you have a theorem universally applicable because it is empty, totally comprehensive, because it expresses simple identity" (41).
The theory of evolutionary change as a consequence of natural selection promoting the adaptation of organisms to their environments is empirically demonstrable in the laboratory and in nature (see I.3.2). However, the difficulties involved in defining adaptation operationally and the subtle relationship between adaptation and fitness cast doubts on the integrity of the theory of natural selection as an all-inclusive theory to account for the origin of life. The fact is that the evolution of life from a single origin, an assertion adamantly maintained by most evolutionists, is more an a priori assumption than an empirically falsifiable theory.
The attempts to analyze the rate of molecular evolution by comparing amino acid sequences of proteins and determining the adaptive values of protein polymorphisms in order to illuminate the neutralist-selectionist controversy will yield nothing more than circumstantial evidence that will be subject to reinterpretation. No one can design any experiment or collect any amount of data from nature to falsify the claim that organic evolution has occurred. The legitimacy of extrapolating microevolutionary observations to macroevolution is increasingly being questioned (27, 28, 33, 38, 42).
The empirical unfalsifiability of the theory of organic evolution has removed it effectively from the realm of empirical science. It is apparent that theories of origins go beyond the limitations of verifiable empirical science and thus require philosophical assumptions and leaps of faith. We will now examine an alternative to the Darwinian theory based on a Christian theistic world view.
![]()
![]()
![]()
References 3.3