 |
|
IBRI Research Report #50 (2001)
Copyright © 2001 by David C, Bossard. All
rights
reserved.
ABSTRACT
| The laws of physics and chemistry are so right that they seem to be designed to support life. Water, carbon, nitrogen and other elements have just the right properties. Subtle electronic forces are able to carry out the life functions spontaneously, given rightly configured molecules. The temperature is right. Creation is fit to support a living cell. Now on to the actual building of life. In this talk, we look at the construction plan for all living cells to see how it is done. Why is it done this way? Is another way possible (yes!)? How hard would it be to build another kind of life? By considering these matters, we can gain an appreciation of how complex the task of creating life is, and begin to face the problems inherent in the view that life is a random product of undirected natural causes. |
EDITOR'S NOTE
| Although the author is in agreement with the doctrinal statement of IBRI, it does not follow that all of the viewpoints espoused in this paper represent official positions of IBRI. Since one of the purposes of the IBRI report series is to serve as a preprint forum, it is possible that the author has revised some aspects of this work since it was first written. |
The Chemical Building Blocks of Life
| “He described plant life, from the cedar of Lebanon to the
hyssop that
grows out of walls. He also taught about animals and birds, reptiles
and
fish.”
I Kings 4:33, NIV
|
This is one in a series of talks that probes the question, "what is life?" This talk concerns the building blocks of life.
The first talk, some time ago, concerns the creation of the elements1, and notes that one cannot just create any old elements in any old universe. There are a number of very precise things that must happen in cosmology and physics so that life can occur. This is the essence of the Anthropic principle, the subject of a number of investigations since the 1950’s when Nobel physicist Fred Hoyle marveled at the fine-tuning of certain physical constants. Later, Hoyle remarked:
A commonsense interpretation of the facts suggests that a super intellect has monkeyed with physics, as well as chemistry and biology, and that there are no blind forces worth speaking about in nature.A later talk concerns “The Right Chemistry for Life”3. This talk notes the fact that water and carbon, together with a few other elements, but especiallythese, are absolutely essential to have life of any sort, whether on this earth or on some alien abode elsewhere in the universe. Don't misunderstand: not just "life as we know it" but "life of any sort." Well, maybe I should hedge a bit: I mean any kind of advanced multi-cellular life, or as someone said, "any life able to pose the question, 'what is life?' "4Fred Hoyle2
So there is water—water, water everywhere. Life is saturated with water, inside and out. And there is carbon to make the stringy molecules called carbon chains. There simply is no other possibility. Liquid ammonia, for example, doesn’t support life, or sulphur or silicon in place of oxygen or carbon5. That's the point of the last talk, and we won't revisit the arguments here.
Besides water and carbon there are a few other elements that are needed for any kind of life. Nitrogen, phosphorous, sulphur, and a dozen or so other elements figure prominently in the cell processes mentioned here. We will not argue for their necessity, just use them as they are appropriate. All of the needed elements are available in the atmosphere or in the minerals that make up the earth's surface, and all occur in forms that are soluble in water—which is good, because that is how they are transported to a cell.
Now the question before us is this: ok, you have water, carbon and these other ingredients. Now what do you need to do to start building life?
We will proceed as follows. First we will survey all of life, to see how many varieties of life exist. Next we will look at the most basic feature that is found in all of these various species, the genetic process called the “central dogma.” Thirdly we will look at a number of representative species from the simplest to most complex to see how much genetic code is needed to specify these species.
After this survey of the data we will look at three questions:
• How “simple” is the smallest, presumably simplest living species that can be imagined?It may be useful to begin by sharing my personal views about evolution. I believe that God created a universe designed to operate in an orderly way according to carefully designed natural laws. This design includes the ability of living species to make genetic changes over time, so that they can survive in varied and changing environments.
• Are other radically different forms of life possible?
• Is it plausible to hold the view that life is a random product of undirected natural causes? What kind of experimental demonstration is needed to support such a view?
It is legitimate science to try to identify the laws of such genetic change, and by implication to try to find relationships among the many species of life. The proper approach to such study includes the systematic description of life as we see it, the postulating of relationships among these species, and the design of repeatable experiments and statistical methods to verify these proposed relationships.
It is not improper to hypothesize that life arose from nonliving matter by random undirected natural processes, but given that hypothesis, the burden is on those who propose it to also propose testable ways to demonstrate its validity. It is not proper science to insist on such a hypothesis, in the absence of such demonstration, as an article of faith that every “proper” scientist must accept.
The Bible talks about God making “kinds” of plants and animals. I believe that this is an accurate statement of God’s activity in Creation, but I do not know the boundaries of these “kinds” and what species are included in each. Presumably all species that are members of a “kind” are related in some way, and I consider the probe for such relationships as good science, as long as the conclusions are based on repeatable experiments, and valid statistical methods.6
Furthermore I do not know the way that God carried out his various creative activities, whether he did them immediately, that is, by fiat, or mediately, that is, using natural laws or means. When existing laws and means were used, I believe that careful scientific investigation will very likely reveal that fact. I believe this because the recent record of scientific findings in physics and chemistry shows that God tends to leave traces of his activity that can be revealed by careful study.
So I welcome the search for relationships among the living species, and for natural mechanisms which produce changes in living forms, and this to me is what evolution science should be all about. Recent years have vastly expanded the capability to study these relationships more intelligently and I eagerly welcome that study, which has barely begun. My only caveat is that claims must use lines of argument that come from objective repeatable experiments and solid deductive reasoning.
Having said this, I do not believe that the evidence to date supports the hypothesis that life arose from nonliving matter by random undirected natural processes, or that all living species are related by natural descent. But I do believe that there are demonstrable relationships out there to be identified, and I suspect that some of these demonstrations will be very surprising and unexpected. It is the anticipation of such surprises that makes me reluctant to dismiss any particular species relationships out of hand. Anything is possible—but show me! The continuing discovery of these relationships is very exciting, rewarding, and quite legitimate science.
I doubt that God created a world with the self-generating powers that some secular—and even some Christian—scientists imagine. I don’t think God gave nature such God-like powers any more than he gave humans more of himself than his image. As it is, the natural world’s amazing capabilities have turned some heads to worship the creature rather than the Creator.
With this said, let us begin.
1. The Three Domains of Life7
Within my lifetime the accepted way to group living matter has changed twice. Early on, I was told that life falls into two kingdoms: plants and animals. This division was a bit subjective and sometimes led to endless and perhaps fruitless discussions about whether such-and-such is a plant or an animal. I sometimes got caught up in that myself.
And so some biologists devised an objective way to look at living species based on microscopic cell characteristics. In this scheme, all of life is divided into prokaryotes and eukaryotes. Eukaryotes have cells with an internal structure called the cytoskeleton, that supports a proper nucleus and other internal organelles. Prokaryotes are everything else.
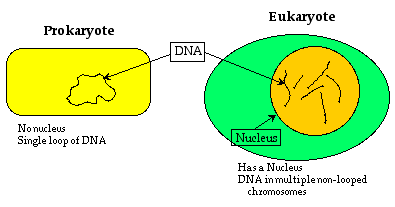
Although the nucleus is quite visible, the supporting cytoskeleton is not. There are three kinds of cytoskeleton filaments, each with its own function: the microfilaments, microtubules and intermediate filaments. The filaments are so fine that they are not normally visible in a microscope, but they can be seen by attaching flourescent dye markers to makes them visible; see for example the illustrations in a 1998 Scientific American article, the “Architecture of Life”.8 Only recently have the details of the eukaryote’s cytoskeleton been revealed.
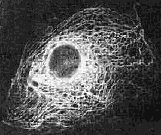
Figure 2: Eukaryote nucleus and cytoskeleton
prepared with flourescent dye.9
The step from prokaryotes to eukaryotes marks a vast increase in cell complexity, reflected (as we will see) in the size of the genetic code. The step also marks a vast increase in capability. In my view the jump in complexity from a prokaryote to a eukaryote is comparable to the jump from non-life to a prokaryote.
To give just one example of this jump in complexity10: Eukaryotes have a highly developed transport system to move food and waste around the cell.11 One component of the cytoskeleton, the microtubules, forms highways, along which special transport molecules called kinesin move, carrying along the food and waste from source to destination, and using a complex dispatching system to keep track of things.
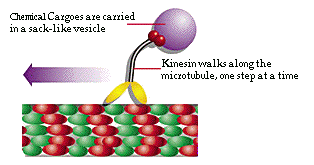
Figure 3: Kinesin Motor12
In contrast, prokaryotes have no transport structure: molecules move about within a cell by ordinary diffusion, the way a drop of dye spreads through water. This is only effective if the cell size is very small, and so prokaryotes are all very small. Eukaryotes, on the other hand, can be large, because they have an efficient transport mechanism. Single nerve cells, for example, can extend many feet.
This division between prokaryotes and eukaryotes lasted until the 1970's when some very strange life forms were found—species that live in "extreme" environments such as around hot vents in the deep ocean, in deep rocks, or around geysers in Yellowstone Park. It was thought at first that these species were heavily adapted prokaryotes that had evolved to survive under harsh conditions. But that view did not survive what happened next.
About the same time a new way arose to look at relationships between species. Historically, the "tree of life" was built by looking at the morphology—the appearance—of different species. That is what you usually see and hear described in the various museums of natural history. But in the 1970's it became possible to study the detailed structure of the genetic code by a process called sequencing. Species could be compared by looking directly at the genetic code. Species that are very closely related have genetic material that looks very similar, and the similarity decreases the more remote the relationship. This is the basis of DNA testing that is done in criminal labs today.
When these methods were applied to the different life species, some surprises occurred. I remember reading in the 1970's that this way of determining relationships was upsetting some assumed relationships of traditional biology, and wondering at the time where it all would lead.
When sequencing13 was applied to the strange creatures of extreme environments, and as more and more species of them were found, they appeared to be closer to the eukaryotes than to the prokaryotes, and they did not appear to be adapted from prokaryotes. In the final analysis, they differ so much from other life that they were called a third major form of life, the Archaea. This divides the prokaryotes into two domains, the Bacteria or Eu-bacteria (“proper” bacteria), and the Archaea or Archaeobacteria (“old” bacteria). The eukaryotes are all in the domain Eukarya.
Another thing happened. Because they thrive in extreme environments—very hot, very salty, highly acidic, etc.—it was thought for a while that the Archaeobacteria were carryovers from the earliest living matter—which accounts for the name "Archaea" —because the earliest life forms must have had to cope with similarly harsh environments. But the genetic studies did not support this: the extreme species are too complex to be that primitive.
Because of the lack of clear evidence of ancestry, the modern way to order these domains avoids putting the Archaea at the bottom—in fact none of the domains is at the bottom. The relationship is usually pictured something like this:
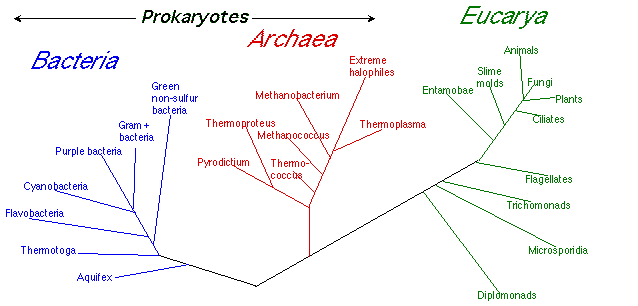
Figure 4: The Modern Domains of Life14
No domain is viewed as the ancestor of—or more "primitive" than—another domain (although eukaryotes are more complex than prokaryotes). The ancestors are pooled in some as-yet-unidentified collection of early life forms, sometimes called “ur-biotica”. There is no living example of a species of ur-biotica.
Not everyone is happy with the current classification scheme. The evolutionist’s desire to classify species in a way that shows a clear path of evolutionary advancement from the earliest life to the present has so far been a failure. The picture is so complex and confused that it has been remarked:
"Microbiology is a field that has developed from its inception virtually untouched by Darwin's grand ideas.... We do not understand how one bacterial phenotype might have arisen from another. We tend not to take evolutionary considerations into account in any discussion…."15As I see it, a reason for this lack of “touch” with Darwinism is that microbiology gets closer to the true nature of biological relationships and the complexity of these relationships makes it harder to imagine the same smooth transitions that were assumed when the comparison was based primarily on physical appearance (morphology) or the fossil record.
A recent article in Sciencemagazine sums this situation up with the provocative title, "Is It Time to Uproot the Tree of Life?". It calls the current classification schemes "a study in confusion."16
2. The Common Feature of All Life: The Central Dogma.
We want to look at life and see how it works. To keep things simple, we will concentrate on so-called “free-living cells”, that is, the single-cell species included in the tree of life. We will look at only those things that are common to all of life.
At a minimum, a free-living cell must keep itself going. This means that it must make the necessary molecules to grow and reproduce. Along the way it must also defend itself against various threats, repair damage to itself, gather food and energy, and dispose of waste products. And it must be able to adapt to a new or changing environment—or else it will go extinct. One outline of these tasks of life lists some 130 functional categories grouped under about a dozen headings.17
Everything that goes on in a cell is controlled by the cell’s genetic code. The genetic code defines and produces the carbon chain molecules that carry out all life functions. The way that the cell does this is called the central dogma. It was first described in the 1950's. It is very detailed and very complex, and it is the same for all of life.
The outline of the central dogma is this:
• DNA is transcribed intoDNA is the code for everything that defines a “species.” DNA is short for deoxyribonucleic acid. It is made up of hundreds or thousands of pieces of code called genes.Each gene gives the blueprint for a particular organic molecule.
• RNA which is translated into
• Proteins which by enzyme action make the
• Carbon chains needed to carry out the life processes.
The DNA does not form carbon molecules by itself. The first step in this process is that the DNA coding for genes is copied into another type of molecule called ribonucleic acid, RNA.
Some of the RNA molecules carry the gene code for specialized molecules called proteins. Some of these proteins in turn are used as enzymes to make the carbon molecules of life.
3. How Does the Central Dogma Work?18
Let us now look more carefully at the central dogma, since it is clearly key to the whole concept of life as we know it.
The DNA holds the genetic code, which is unique for each species. Four carbon chain molecules called nucleotides, form the genetic code. The letters G,C,A and T represent the four nucleotides19. They always appear in DNA as pairs G-C and A-T, called "base pairs". These same molecules appear in every living species.
DNA is a long molecule built like a spiral ladder with a right-handed twist. The ladder sides are chains of sugar and phosphate, and the "rungs" are the base pairs. The sequence of the pairs determines the genetic code. Every protein, RNA or other molecule manufactured in a living cell has its own defining code sequence recorded in the DNA.
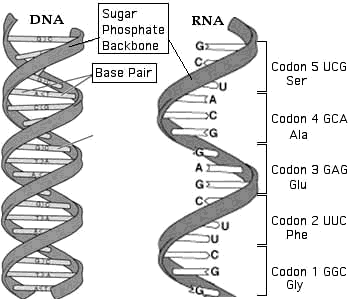
Figure 5: DNA and RNA Molecules
RNA is obtained from DNA by a copying process called transcription. All species produce RNA in this way: A special type of RNA molecule called RNA polymerase moves along the DNA and causes it to unwind by temporarily breaking the G-C and A-T pairs. One side of the DNA becomes a template that is used to build an RNA molecule that is identical to the missing side of the DNA, with T replaced by a slightly different nucleotide, U.
Many RNA molecules may be produced simultaneously along a given DNA gene segment. The following photograph shows this transcription process in action.
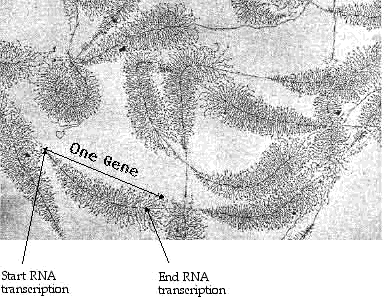
Figure 6: RNA Transcription from DNA20
In this figure, several distinct DNA genes are visible, most of them undergoing transcription. The fern-like appearance is caused by many RNA copies of each gene made at the same time. Copying begins at the start of the gene—the narrow tip of the "fern" with the short, newly formed RNA—and ends at the base of the "fern", with the RNA at full length. The numerous RNA polymerase molecules are visible as a darkening of the faint DNA trace. The DNA would be seen as a single long snarled strand.
An RNA molecule that codes for a protein is called messenger RNA, denoted mRNA. When transcription from the DNA is complete, the mRNA molecule moves away from the DNA.
Proteins are made up of small carbon chain molecules called amino acids21. There are 20 different amino acids that can be specified by the DNA code. For example, here is a model of glycine, the simplest amino acid:
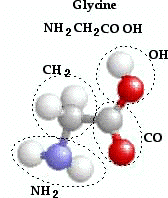
Figure 7: Glycine, the simplest Amino Acid22
The twenty amino acids are usually designated by a 3-letter abbreviation, such as “gly” for glycine, etc.
The genetic code transcribed from DNA into the mRNA specifies the exact sequence of these amino acids in the protein molecule. The code is read in groups of three, called “codons.” The code may be read in a forward direction (up the spiral, as shown in Figure 5) or in the reverse direction.
Every possible combination of 3 nucleotides represents a specific amino acid, or else a coding instruction. For example, the codes AGA and AGG both represent Arg=arginine and the codes UAA and UAG are “stop” codons. Figure 5 shows some of these codons.
All living species use the same triplet codon scheme to specify the amino acids, with only some minor variations in the exact meaning of a given codon23.
Special RNA molecules, called transfer RNA, tRNA, read the codons and translate them into specific amino acids. A separate tRNA molecule exists for most distinct codons24. Of course each tRNA molecule must be built by the cell, and its code included in the DNA. Every species of life requires dozens of different tRNA molecules to translate the 64 possible codons into the twenty amino acids.25
The stop codon does not specify a particular amino acid; instead, it corresponds to a special molecule called the “release factor” which ends the protein building process and releases the protein from the rRNA.
The proteins are formed as a growing chain of amino acids, by a very complex molecule called ribosomal RNA, rRNA. Typically, rRNA is a complex of 50 or more separate protein and RNA molecules. The ribosome forms the protein chain by moving along the mRNA, growing the protein chain as it proceeds. This is called RNA translation (to distinguish it from DNA transcription).26 A photograph showing ribosomes building proteins along the length of an mRNA molecule can be seen as part of a Drake University lecture.27
4. Enzymes.
The central dogma would not work without the help of many enzymes. Enzymes are special molecules (usually proteins) that help many chemical reactions to take place. Without the enzymes the reactions would not occur at all, or they would be much too slow to keep pace with the needs of the cell.
All carbon chain molecules that carry out the central dogma are built using enzymes. This includes the nucleotides, the RNA polymerase, the amino acids, the ribosomes, the tRNA, and many other specialized molecules that are needed to carry out the central dogma. If a cell needs a particular molecule, it must either obtain it as food (which means some other living cell produced it using enzymes), or it must include the DNA code for that molecule plus coding for all the enzymes needed to build it.
Enzymes hold molecules in a particular way so that certain reactions can take place, as illustrated in Figure 8. In this figure, the enzyme joins two pieces of food, called the “substrate”. Enzymes can also be used to break molecules into pieces, for example to break down food into useable parts. These are called “cutting enzymes”.
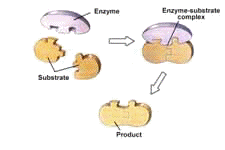
Figure 8: Enzyme Promoting Molecule Formation28
To picture this enzyme activity, think of threading a needle. If the thread and needle are both floating in mid-air, the thread may occasionally float through the needle’s eye, but that would not happen very often. In the same way, a reaction without the help of an enzyme may occasionally happen, but it will not happen often enough to be of value.29
To get the needle threaded, it is necessary to hold both the needle and thread in just the right way, and then move one towards the other just so, and the job is done. The hands in this illustration are like enzyme molecules holding the separate parts, and the motion is like the subtle electronic forces that pull the parts together to make the product. Without the enzymes and the subtle forces, the job will not get done effectively.
An enzyme has a shape that attracts a particular kind of molecule, and that is why the right kinds of molecules are brought together. This use of reaction sites is basic to the whole fabric of the central dogma.
Without enzymes it would be very hard to manufacture the molecules needed by a cell. Some complex molecules require the assistance of dozens of different enzymes in their manufacture. The building sequence is called the metabolic pathway. Many of these metabolic pathways are the same or very similar across nearly all species of life and are given names, such as the citric acid cycle, the glycolysis cycle, etc.
5. The size of the genetic code.
The point of this survey of the central dogma is that every living cell carries out the same complex process so that it can make the molecules that it needs to carry out its life functions. There is no living species in any of the three domains of life that carries out, say, only half of the central dogma. You might well wonder why all species use such an indirect way to produce the molecules needed for life. Why, for example, isn’t there some species that uses a “primitive” version of the central dogma in which the DNA makes the end products itself instead of using the indirect RNA route? The only apparent shortcut to this complicated process is to use organic food prepared by some other species. For example, humans need certain vitamins that their own cells do not make in suffient amounts.
Every living cell of every known species in any of the three domains
of life—Bacteria, Archaea or Eukarya—has to do the things we have
talked
about, and much more. This takes a lot of genetic code. How much? In
the
past few years, scientists have sequenced the complete genetic code for
a number of species. The following table shows the size of this code
for
a few examples that are of special interest. The size is given in terms
of the number of base pairs—rungs in the DNA ladder—and the number of
genes.
|
|
|||
| Species (year sequenced)32 |
Base pairs | Genes | Notes |
| Mycoplasma genitalium (1995)33 |
|
|
The smallest
species DNA known to date. Bacteria Domain. Lives (harmlessly) in the genital tract. |
| Methanobacterium
thermoautotrophicum (1998)34 |
|
|
Smallest
autotroph DNA known to date (see text). Archaea Domain. |
| Synechocystis (1996) |
|
|
a marine
photosynthetic cyanobacteria ("blue-green algae"). Bacteria Domain. The oldest known fossils on Earth appear similar to this species. |
| Escherichia coli (1997)35 |
|
|
Bacteria domain.
Resides in the human digestive tract. |
| Saccharomyces cerevisiae (1996) |
|
|
Budding yeast.
The smallest eukaryote
DNA known to date. Eukarya Domain. Includes 275 tRNA and 140 rRNA genes. |
| Arabidopsis
thaliana (December, 2000)36 |
|
|
Mustard family.
Smallest plant DNA
known to date. Eukarya Domain. |
| Humans (in process)37 | 3,446,197,000 38 | ~30,00039 | Exact gene count will be known soon. |
The first table entry, Mycoplasma genitalium, was the first species to have its genetic code completely deciphered. This was done in 1995. It has the smallest genetic code of any species. Mycoplasma genitalium is a harmless bacterium that lives in the human genital tract (hence the name).
Note that the "simplest" living species in the world today needs nearly 500 genes and over 500,000 base pairs in its genetic code. Of these 500 genes, about 152 genes are needed just to carry out the central dogma; 55 genes are needed just to build the single ribosome that the cell needs to manufacture proteins from the genetic code; 24 genes build the tRNA; 21 genes are used to make amino acids, and so on.30
M. genitalium can get away with so few genes because it depends on a rich supply of organic food for many of the molecules that it needs to survive. In contrast, the next entry, M. thermoautotrophicum, is the smallest known species that is an autotroph, that is, it can manufacture all of its own food from basic minerals and simple chemical compounds. It is a methane-producing microbe from the domain Archaea.
One can easily imagine that the very first species on earth would have to be autotrophs, because there would not be any other living species to provide their food. But the size of the genetic code is a problem. In order to survive without organic food, the autotrophs require over 2000 genes and 1.7 million base pairs, several times that of Mycoplasma.
The next species in the table, Synechocystis, presents us with another problem. It is from the phylum Cyanobacteria, sometimes called blue-green algae. It has chlorophyll that can convert sunlight into energy and use that energy to make food in the form of sugar. Note that it has over twice the number of base pairs and genes as M. thermoautotrophicum. These cyanobacteria are single-celled, but the cells reproduce by dividing into halves that remain attached at the ends.
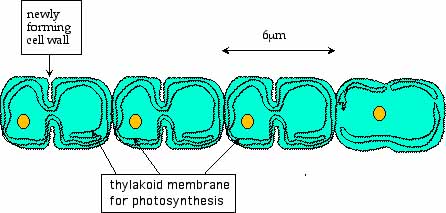
Figure 9: Modern Cyanobacteria40
Here is the problem: The very oldest fossils found on earth look
like
shrivelled, dried out chains of Cyanobacteria. They look something like
the following sketch:
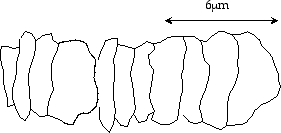
Figure 10: Typical fossil cyanobacteria
similar to the oldest fossils on earth42
These fossils come from a time that is just after the earth first cooled to the point where liquid oceans could exist. How could such “advanced” species have evolved in so short a time—a time that is so short it is less than the measurement error for early earth dates? One might as well assume that life arose instantaneously41.
Of course we do not have the actual genetic material from these first fossils, but it doesn’t seem likely that they are much simpler than modern cyanobacteria, which have an exceedingly complex genetic code, almost twice a large as M. thermoautrophicum!
The listing for E. coli is included because it is the most studied bacterium in all of biology. It is found in everyone's digestive tract, in thousands of varieties, mostly beneficial or harmless, but some subspecies are harmful.
The next entry in the table, Saccharomyces cerevisiae, is the eukaryote with the smallest known genetic code. As we noted earlier, eukaryotes have a cytoskeleton that gives an orderly structure to the cell and supports an internal transit system. These "innovations" are needed to allow the formation of large (compared with bacteria and archaea), complex, highly structured cells and multi-cellular bodies, and is the reason why all higher species including plants and animals are eukaryotes. All such species have DNA with over 12 million base pairs and 5800 genes.
Arabidopsis thaliana has the smallest DNA of any flowering plant. Note another jump in complexity, to 120 million base pairs and about 25000 genes.
A. thaliana is one of the wild mustards familiar as a weed to American gardeners and grows throughout Europe and Asia as well. It is closely related to other mustards: the common mustard that is seen as a yellow-flowered weed that grows in fallow fields throughout America, and the black mustard that Jesus called “the smallest seed” in Matthew 13:32. In my imagination I picture Jesus making this remark as he walked by a fallow field in Palestine, a field covered with mustard weeds that can reach to twice a man’s height.

Common Mustard
Since the mustard is the plant with the smallest genetic code, perhaps Jesus’ remark is accurate in a deeper sense than many Bible scholars may have supposed.43 It seems particularly interesting that the author of Matthew follows the parable of the mustard seed with the remark that in using parables, Jesus said “things hidden since the creation of the world.” [Matthew 13:35b, NIV].
Finally, I added Humans, to indicate the size of human genetic code in comparison with the other species in the table.
6. Search for the Simplest Possible Life.
The extreme complexity of all living species should be a bit unsettling to those who hold to undirected natural evolution. The question naturally arises whether this complexity is necessary, or whether it just reflects the complicated fine-tuning of billions of years of evolution. In 1998 the National Research Council of the National Academy of Science held a symposium to look at theoretical lower limits to the complexity of life. The symposium had the title, Size Limits of Very Small Microorganisms.44 The report was published in 2000.
The symposium looked at size limits dictated by physical and chemical as well as biological considerations, so it was a genuine cross-discipline effort.
There are two answers to the question of size limits, depending on the kind of food that is available. If the cell can use organic food, that is, food that is prepared by another living cell, then the DNA doesn't have to manufacture all of its own life molecules. Thus M. genitalium only makes one of its twenty amino acids, relying for protein food to provide the others. On the other hand, if the cell has to make all of its own food from simple inorganic material, then the DNA has to include the genes needed to manufacture all of the amino acids, as well as all other life molecules.
The Symposium concentrated on the simpler case. Its conclusion is
that
"Free-living organisms require a minumum of 250-450 proteins along with the genes and ribosomes necessary for their synthesis. [A sphere having a diameter of] 250 ± 50 nm constitutes a reasonable lower size limit for life as we know it."45
M. genitalium is not very far from the simplest possible life form
that can be imagined. One symposium participant stated:
"Active growing cells significantly smaller than those of M. genitalium are unlikely to exist on Earth today. In addition, the limitations on genome size that accompany small cell size guarantee that cells of that size will have to depend on larger, more complex organisms for supplies of most of the small organic molecules required for their growth. Hence, a biosphere consisting entirely of ultra-small organisms is highly implausible."46
I should say something about a recent event that was a factor that
led to the symposium. In August 1996 NASA announced possible evidence
of
past life on Mars. A meteorite from Mars was found in Antartica and
seemed
to show microbial fossils.
Some prominent scientists, including J. William Schopf, questioned the finding. An account of his involvement in the aftermath of the announcement is described in the last chapter of his recent book, Cradle of Life.47 Schopf questioned whether the objects found by NASA were fossils because they were far too small to hold enough molecules to support life. He remarked:
“Their 20 to 30 nm breadth is thinner even than the simplest bacterial cell wall…the living cavity would have been miniscule.…For the “Mars microbes” to grow and reproduce…each of their biochemicals would have to do the work of some two thousand earthly counterpart molecules. Something is amiss. How could such tiny microbes live?”48He illustrates in a diagram49 that the “Martian microbes” would have a volume 2000 times smaller than M. genitalium, the smallest species on earth.
The conclusion of the symposium would seem to rule out the “Martian microbes” as fossils of once living matter.
7. “Contingency” and new kinds of life
So here we are. We have just spent some time looking closely at the way that every viable cell, from the simplest to the most complex, carries out the central dogma. We have looked at the empirical evidence about the size and complexity of all living species. We have seen the results of a symposium on the Size Limits of Very Small Microorganisms. All of which clouds the question of the origin of life to a person who believes that life arose by undirected natural evolution.
The overall impression that you must surely get is that life, at least the only life we can observe at the present, is incredibly complex. Even the most basic process that sets up the production of the molecules needed to carry out the simplest cell's activities, is so complex it is hard to imagine how it could arise by undirected random processes. Hundreds of genes involving hundreds of thousands of base pairs are needed just to carry out the Central Dogma. Hundreds more genes are needed to carry on the cell's life functions. Ironically, even the production of the components of the central dogma depend on the whole transcription process being in place for their own production. How, one might ask, does one get started?
But perhaps this apparent complexity is misleading. Perhaps, far from being complex, the life we see on Earth is inevitable! That is what some biologists such as Stephen Jay Gould assert.
To say life as we know it is inevitable and yet to know that there is only one kind of life is, in effect, to say the appearance of contingency is an illusion. Does this make sense?
Contingency is the idea that certain fixed features of life appear to be selected (either randomly or by choice) from a range of possibilities. Webster says that a feature is contingent if it is not the only possibility—it is “possible but not certain.”
If you chose a cracker from a plate of animal crackers made in a variety of shapes, you made a contingent choice, because you picked a particular design, from a number of equally valid possibilities. This is true whether you made a deliberate choice or just picked one at random.
There are many apparent contingencies to be found in life as we know it, at least as far as scientists can determine at present. For example:
• Could other molecules substitute for the 4 nucleotides?And there are others that we haven’t mentioned today:
• Could other amino acids substitute for the 20 amino acids? Why only 20?
• Could other elements substitute for nitrogen, phosphorous, and sulfur?
• Is the codon triplet scheme the only possible coding scheme?
• Is the 4-stage Central Dogma the only building plan possible?
• The fact that DNA and all of the nucleotides have right-hand twists whereas all of the amino acids have left-hand twists.The list could go on and on. What is there about life that requires these specific choices as the only ones used by all of life? The answer, as far as we know, is “nothing.” In other words all of these choices have the appearance of being contingent.
• The use of a particular phosphorous-based molecule called ATP as a universal “battery” for energy storage and retrieval.
• The universal use of a rare metal, molybdenum, in the enzyme nitrogenase, used to make nitrogen available for use by life processes50.
• The use of the chlorophyll molecules to convert light to energy.
It would seem that one could create entirely new forms of life (at least, in principle, although it would be an exceedingly difficult job!) by making some small changes in a few of these apparent contingencies. For example, right-handed proteins have been made in the laboratory: in principle, why couldn’t an entire living species be made with right-handed proteins? There is no known physical reason why this could not be done.51The result would be an entirely new kind of life.
Or, perhaps the complex central dogma could be much simpler. Again the result would be an entirely new kind of life. My response to this is, "I am willing to be shown: show me."
8. Challenges for Natural Evolution
All of this gives some real problems to those who believe that life is a random product of undirected natural causes. Here are some of the problems that such a person faces:
• How did life arise so quickly?If evolution makes any sense at all, then it must be based on a gradual increase in complexity. Any gradual scenario is seriously compromised by the apparent rapidity with which life first appeared on earth.
• Why did life arise only once?The universal conclusion of biologists that life arose only once comes from the fact that many universal features of life appear to be contingent, which makes it implausible that exactly the same kind of life arose more than once.
Certainly life appearing all at once in all of its glory—even the lowly “simple” prokaryote life—is not evolution by any stretch of the imagination. An ad-hocexplanation might be that life is inevitable, given a hospitable environment. That is what Stephen Jay Gould asserted. But in that case why did it arise only once—or if it arose more than once how is it possible that every line but the one we have today disappeared without a trace?
• What are viable but simplified versions of the central dogma?An evolutionist must believe that radically simplified versions of the central dogma are possible. There are no examples around to point to, but they must have existed in evolutionary history. What are they? Even if the Earth environment has changed, the chemical and physical laws have not, so it must be possible to produce viable intermediate stages of life all along the line. If they survived to produce the current species, then they must be able to survive long enough to be demonstrated in a laboratory. I am not saying that this is easy, or even possible with today’s tools, but surely testable theoretical models must be out there to find.
As far as I am aware, the testable hypotheses that are needed to prove the validity of evolution are notably missing. This is not to say that hypotheses have not been offered from time to time, but as far as I know they have not fared very well under close scrutiny, or else they have raised as many questions as they answered.
9. Conclusion
I would like to conclude with a summary of what we have discussed.
• No “simple” life form exists.“Simple” life is a fantasy of wishful thinking. There is nothing simple about even the lowliest living species. The word should be avoided in all discussion of life, because it implies something that does not exist. Similarly, the “pro” in prokaryote and “archaeo” in archeobacteria are misnomers, because they both imply an ancestry relationship that the evidence does not support.
• Life substantially smaller or simpler than the smallest species found today does not appear possible, as viewed by physics, chemistry or genetics.There is a lower limit on size and complexity, based not just on the observation of existing species, but on theoretical grounds as well. That lower limit is closely approached by the living species M. genitalium which has over 500 genes and a DNA with over 500,000 base pairs. Even to reach this minimum, it is necessary to take substantial short cuts that require a rich organic environment to survive.
• No clear ancestry relationship exists among the major domains of life.There are (at least) three basic plans represented by the Bacteria, Archaea and Eukarya domains, but there is no clear indication that any domain is an ancestor of any other.
• All living species follow the same complex Central Dogma to build life molecules.There is no known practical or theoretical way to construct a viable life form that has a significantly simplified version of the central dogma.
• Life appeared on Earth as soon as the physical conditions could support it.The earliest evidence of life is recognizably similar to a modern photosynthetic species. The modern species has a complex genetic makeup and there is no evidence that the ancient species were substantially simpler.
• Many features of life appear to be “contingent” and yet they are the same across all species.Every known species of life shares the same exceedingly complex building plan, even though this plan appears to have a number of features that do not appear to be pre-determined by the natural laws.
One possible conclusion is that there is no contingency—it is all just an illusion. But if this is so then there must be some unknown natural laws that lead to a directed result. It is the evolutionist’s burden to identify these unknown laws and demonstrate them through effective experiments.
On the other hand if there is only a single original life form, then the burden of the evolutionist is to explain the relationships between the domains of life, and to suggest testable hypotheses to show how the divergences arose.
As I see it, the evidence we have looked at here amounts to almost overwhelming obstacles that a person who believes in undirected natural evolution must overcome. At the very least, there is a lot that needs to be proved and demonstrated.
As one who believes that God created life these features of life are confirming evidence of God’s activity and show that life is the product of wisdom and intelligent design. Common features of species may or may not indicate common ancestry—an equally good possibility is that they indicate a common Creator.
Let’s end by coming full circle: I agree with Hoyle’s commonsense interpretation of the facts: a super intellect has monkeyed with the physics, chemistry and biology. There are no blind forces worth speaking about in nature.
FOOTNOTES
1 David C. Bossard,
A
Physicist Looks at Creation, IBRI Research Report #49.
Available
at the IBRI website, http://www.ibri.org/.
2 Fred Hoyle, Steady-State Cosmology Revisited,Cardiff Press, 1980, cited in Fred Hoyle, "The Universe: Past and Present Reflections", Annual Review of Astronomy and Astrophysics,20 (1982), p4. Also quoted in Michael J. Denton, Nature’s Destiny: How the Laws of Biology Reveal Purpose in the Universe,Free Press, 1998, p12.
3 David C. Bossard, The Right Chemistry for Life, IBRI Colloquium Series, March, 2000. Available at the IBRI website, http://www.ibri.org/.
4 Actually, I believe the more general “life of any sort” as this talk will demonstrate. Denton, op. cit.for example. does not rule out the spontaneous generation of “simple” life. Peter D. Ward and Donald Brownlee, Rare Earth,Copernicus, 2000 pxiv states “We are not saying that lifeis rare—only that animallife is.” In my view the argument that “simple” life may be common in the universe is an argument from ignorance—the more we learn about “simple” life, the less simple it appears.
5 So-called sulfur-based life still requires oxygen.
6 According to one author, there is a “species problem” in biology today. Features that distinguish most species are usually trivial and have little to do with adaptation or survival. See Jeffrey H. Schwartz, Sudden Origins: Fossils, Genes and the Emergence of Species,John Wiley & Sons, 1999, p.203. The implication is that a more scientific approach to determining species may yield some surprises.
7 For discussion of this history see W. Ford Doolittle, "What are the archaebacteria..." in M.J.Danson, et al,eds., Archaebacteria: Biochemistry and Biotechnology,Elsevier, 1993, p1. Some authors do not use the Domain division described here. For example, Lynn Margulis divides living species into five kingdoms. See Lynn Margulis and Kathlene V. Schwartz, Five Kingdoms: An Illustrated Guide to the Phyla of Earth,Third Edition, W.H. Freeman, 1999, p79.
8 To see photos of each type of cytoskeleton filament, consult the article published by Donald E. Ingber, “The Architecture of Life”, Scientific American, January 1998. This article is available for purchase online at https://www.sciamdigital.com/ for a nominal fee.
9 http://www.wfu.edu/Academic-departments/Biology/courses/bio214/images/cytoskeleton.htm. (link no longer available)
10 see http://virtual.class.uconn.edu/~terry/Common/3domains.html for a discussion of organelles and other defining features of eukaryotes.
11 See Bruce Alberts, et al, Essential Cell Biology: An Introduction to the Molecular Biology of the Cell,Garland Publishing, 1998, ch. 16 “Cytoskeleton”, p525-7. The “motors” are called kinesin.See http://www.blocks.fhcrc.org/~kinesin/ and Howard Hughes Medical Institute, “Learning How a Cell’s Tiny Motor Powers its Mobility”, 2000 at http://www.hhmi.org/news/vale.html.
12 Image obtained from http://www.newscientist.com/ns/19990619/images/kinesin2.jpg from “The Engine of Creation, New Scientist, 19 June 1999. Used by permission.
13 The differences in the makeup of ribosomes (see below) led to the conclusion discussed here.
14 From Woese,
C.
R., Kandler, O. & Wheelis, M. L. (1990) Proc. Natl. Acad. Sci. USA
87, 4576-4579.
For a more detailed tree see, for example: http://www.bact.wisc.edu/bact303/PaceTree.jpeg/
(link no longer available) or
http://virtual.class.uconn.edu/~terry/images/other/3domains.gif.
Used by permission.
15 Carl R. Woese, "Why study evolutionary relationships among bacteria?" in Karl H. Schleifer & Erko Stackebrandt, Evolution of Prokaryotes,Academic Press, 1985, p1.
16 Science284, 21 May 2000, p1305-1307.
17 See, for
example:
http://www.cric.com/genesequences/methanobacter/function_categories.html
(no longer available),
http://hearing.bwh.harvard.edu/knowngenes3.htm
(no longer available), and
http://www.ncbi.nlm.nih.gov/Complete_Genomes/Minset/fun.html
18 A description of the Central Dogma with many graphic illustrations is found in Bruce Alberts, et.al.Essential Cell Biology,Garland Publishing, 1998. Many illustrations from this book are on the web at accessexcellence.org.
19 G = Guanine, C = Cytosine, A = Adenine, T = Thymine, U = Uracil. All nucleotides have a central ring of 4 carbon and 2 nitrogen atoms. Adenine is Vitamin B4.
20 Photograph from O. L. Miller, Jr. “Visualization of genes in action” in Scientific American,Vol 228, March, 1973, p38. See http://www.esb.utexas.edu/dr325/genweb/images/13-4.gif. Used by permission.
21 All amino acids have an amino group NH2 and a carboxyl group COOH and various side chains. In neutral water (PH = 7), NH2 is ionized to NH3+and COOH is ionized to COO-.
22 View the
amino
acids at http://www.chemie.fu-berlin.de/chemistry/bio/amino-acids.html
Key to Diagram: Hydrogen = White, Oxygen = Red, Carbon
= Gray, Nitrogen = Blue.
23 See Brian Hayes, “The Invention of the Genetic Code”, American Scientist,Jan-Feb 1998. Available on line at http://www.americanscientist.org/issues/Comsci98/compsci9801.html. When the triplet codon scheme was first discovered (in the 1960’s) some scientists remarked on its “inefficiency.” Yet all of life uses the same scheme. It seems that the code scheme is designed to be error tolerant: a base pair error usually codes for an amino acid that is the same or close in function to the intended one.
24 Sometimes a single tRNA translates more than one codon.
25 The standard codon table is shown at http://www.accessexcellence.org/RC/VL/GG/genetic.html and many other web sites. Some species use non-standard codons—still triplets, but with different amino acid translations. See for example, http://prowl.rockefeller.edu/aainfo/gencode.html.
26 See http://www.cvcc.cc.nc.us/programs/art/science/DNA1.htm for graphical illustrations of RNA translation.
27 http://pharmacy.drake.edu/faculty/L_Henderson/Unit5/trans-ribo.jpg(Link is no longer active)
28 From http://www.cvcc.cc.nc.us/programs/art/science/competi.jpg. Used by Permission.
29 The Miller-Urey experiment shows that amino acids will form without the help of enzymes. The question is, whether this method can produce sufficient numbers in a useful and timely manner.
30 See http://www.ncbi.nlm.nih.gov/Complete_Genomes/Mgen/fun.html gives a functional summary of the M. genitalium genes. Also see detailed gene descriptions listed in the table notes.
31 The table is
derived
(with minor changes) from
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/G/GenomeSizes.html.
This site is updated periodically, and should be consulted for the
latest
information.
32 For a list
of species
that have been sequenced, see:
http://www.ebi.ac.uk/genomes/
and
http://www.tigr.org/tdb/mdb/mdbcomplete.html
33 A complete
table
of M.genitalium with the identity and function of each gene is at
http://www.ncbi.nlm.nih.gov/cgi-bin/Complete_Genomes/mgtable
34 A complete
table
of M. thermoautotrophicum with the identity and function of each gene
is
at
http://www.ncbi.nlm.nih.gov/cgi-bin/Entrez/altik?gi=130&db=Genome&from=0&to=0
(also see http://www.cric.com/genesequences/methanobacter/abstract.html)
35 A complete
table
of E. Coli with the identity and function of the known genes is at
ftp://ftp.genome.wisc.edu/pub/analysis/.
(table located in m52orfs.txt)
36 Completion was announced December 13, 2000, the day after this talk was first presented. See Nature 408 14 December 2000. Article at http://www.nature.com/genomics/papers/a_thaliana.html. For gene information see also http://www.tigr.org/tdb/ath1/htmls/ath1.html, http://genome-www.stanford.edu/, and http://www.arabidopsis.org/.
37 A private
company,
Celera, claimed to complete the sequencing in April, 2000. The human
genome
project has a scheduled completion date of 2003. A tabulation of
progress
is at
http://www.sanger.ac.uk/HGP/Humana/stats.shtml
and
http://www.ornl.gov/TechResources/Human_Genome/home.html.
For a list of most-used web sites see: http://www-ls.lanl.gov/HGhotlist.html
38 as of September 18, 2000. See http://www.ornl.gov/TechResources/Human_Genome/project/progress.html.
39 The human body has about 250,000 separate proteins, so each gene must code for an average of 10 separate proteins. This is done by the genes splicing their DNA to produce multiple proteins.
40 For illustrations of Anabaena Cyanobacteria see Lynn Margulis and Kathlene V. Schwartz, Op. Cit.,p79.
41 Some scientists, as Stephen Jay Gould, take this sudden appearance of life to “prove” that life is inevitable, and that it therefore must exist in many places throughout the universe. In Wonderful Life: The Burgess Shale and the Nature of History,W.W.Norton, 1989, p309 he wrote: “I do not, by the way, view the origin of life itself as a chancy or unpredictable event. I suspect that given the composition of early atmosphere and oceans, life’s origin was a chemical necessity. Contingency arises later.”
42 See composite photograph in Schopf, Cradle of Life,op. cit.Fig. 3.4. A similar composite is found at http://www.astrobiology.ucla.edu/ESS116/L15/1515%20Apex%20Chert.jpg
43 All mustards
are
of the family Brassicacaea (also called Crucifera). The Biblical
mustard
is generally thought to be Brassica nigra (Sinapis nigra -- black
mustard)
although there is some dispute about this. See various Bible
dictionaries
and the Merriam-Webster Unabridged Dictionary under the entries
“arabidopsis”,
“mouse-eared cress” and “mustard.” For a sketch of some mustards,
including
A. thaliana taken from John Curtis, British Entomology (1824-1835),
see http://biodiversity.uno.edu/delta/angio/images/cruci08.jpg
and
http://biodiversity.uno.edu/delta/angio/images/cruci02.jpg.
44 Size Limits of Very Small Microorganism,National Research Council, 2000. Text is available at http://www7.nationalacademies.org/ssb/nanomenu.html.
45 NRC, ibid.,Andrew Knoll, "Overview", nm = nanometer (a billionth of a meter).
46 NRC, ibid.Peter B. Moore, "A Biophysical Chemist's thoughts on cell size." Abstract.
47 J. William Schopf, Cradle of Life: The Discovery of Earth’s Earliest Fossils,Princeton U. Press, 1999.
49 J. William Schopf, Cradle of Life.Fig. 12.5 p320.
50 See Harold J. Morowitz, Beginnings of Cellular Life,Yale Press, 1992, p43, “It is required in prokaryotes for dinitrogen fixation and in plants and prokaryotes for nitrogen reduction.”
51 Christopher Wills and Jeffrey Bada, The Spark of Life: Darwin andthe Primeval Soup,Perseus Publishing, 2000, p119 report an experiment that created a protein of 99 right-handed amino acids. They summarize: “there is no apparent biochemical reason why L-amino acids would have been selected over D-amino acids when life began.”
 You
can contact IBRI by e-mail at: webmaster@ibri.org
You
can contact IBRI by e-mail at: webmaster@ibri.org
Last updated: November
18,
2002; Hyperlinks updated 21-Jul-2003
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}