2.3 Evidence from Physical Anthropology
To trace the presumed evolutionary lineage of humans, one has to start with living and fossil members of the superfamily Hominoidea. This group consists of all the great and lesser apes and all humans. Table 2.9 represents a summary of the members of superfamily Hominoidea and their known and related fossils.
Table 2.9. Superfamily Hominoidea.
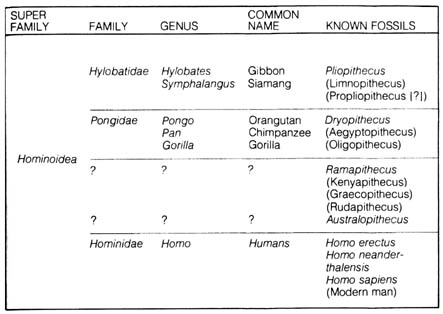
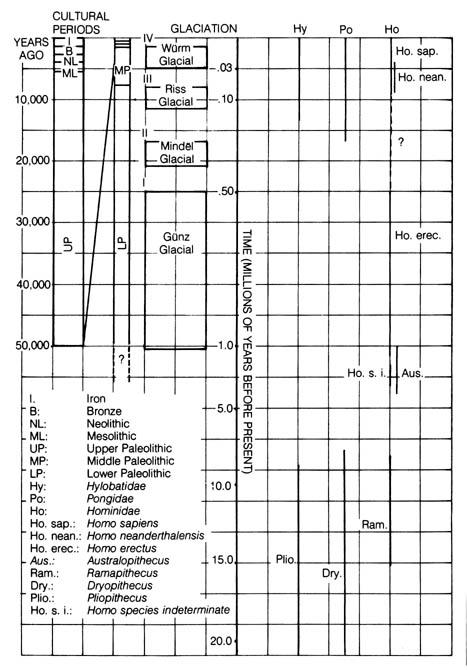
As shown in Figure 2.14, the evolution of Hominoidea can be traced by three separate lines. Each can be represented by an extinct genus, [95] namely, Pliopithecus, Dryopithecus, and Ramapithecus. All three fossil lines were definitely more like apes than humans. However, there have been speculations that Ramapithecus is more humanlike and was the remote ancestor of modern humans (1). Assessment of evolutionary relationships of the fossils above has been based mainly on available anatomical features such as posture, brain size, and dentition (kind, number, and arrangement of the teeth). For example, Table 2.10 summarizes the cranial capacities of several forms within the superfamily Hominoidea.
Table 2.10. Cranial capacities of humans and some presumed relatives.*
Excluding where possible, pathologically abnormal individuals.
* NOTE: Reprinted, with permission, from Lerner, I. M.; Libby, W. J. Heredity, evolution, and society. San Francisco: W. H. Freeman and Co. ©1976.Figure 2.14. Time scale of Hominoidea ancestry as correlated with glaciation and cultural periods.
[97] 2.3.1 Pliopithecus and Dryopithecus. Pliopithecus
was
discovered in Europe from rocks dated between the Miocene (25 million years
ago) and Pliocene (6 million years ago) eras. There were several fossil
finds assigned to this genus, including an almost complete skull. Remains
of a fossil discovered in East Africa in Miocene rocks included parts of
the postcranial skeleton and the limbs. This form was known as
Limnopithecus.
Phopithecus and Limnopithecus are very similar and both seem
to be related more to the family Hylobatidae than to Pongidae
and
Hominidae.
Dryopithecus (2) was found both in Oligocene (38 million years ago) and Miocene rocks and is related to the family Pongidae. The dryopithecines have been divided into four genera, namely, Aegyptopithecus, Oligopithecus, Propliopithecus, and Dryopithecus.
Aegyptopithecus was found in North Africa and was especially important because in addition to five partial lower jaws, an almost complete skull was found. The teeth are low crowned (minimal development of enamel) and not specialized. The jaws protrude beyond the ridge of the upper part of the face, a feature shared by many primitive primates. Members of Aegyptopithecus seem to have made their abode in the forest canopy and probably had a herbivorous (plant-eating) or frugivorous (fruit-eating) diet.
In contrast, Oligopithecus is known only from a single jaw uncovered from an early Oligocene rock. The jaw is comparable in size to that of a squirrel monkey, but the teeth definitely resemble those of higher primates. Although only the lower jaw is available, the pattern of wear in the lower premolar is consistent with that of a higher primate in which the upper canine is commonly honed to a sharp edge against the first lower premolar. Because of the scanty record, the interpretation of the Oligocene evidence is only tentative.
Propliopithecus was found in 1908 in an Oligocene rock in Egypt. The find includes two half mandibles (bone of lower jaw) and some teeth that may or may not have come from the same skeleton. The dentition, with small canines comparable in size and form to the premolars and the low-crowned molars, has caused some anthropologists to link this fossil to the family Hominidae. However, its homology to the modern gibbon is more striking, and it seems more accurate to put Prophopithecus in the lineage of Hylobatidae.
The Miocene dryopithecines are abundant and widely distributed. Dryopithecus was found in Europe, Asia, and Africa and is represented by a large collection of jaws and teeth. This form is also known from Pliocene [98] deposits. The dentition of Dryopithecus is more like that of modern apes than the other finds in the same period.
The African dryopithecine, Dryopithecus africanus (originally named Proconsul), left considerable remains, including limb bones, a skeleton of a hand, and an almost complete skull. There are three species ranging in size from a large gibbon through a chimpanzee to a small gorilla. As evidenced from the limb bones, these Miocene apes seemed to be of a lighter build than modern apes. They were most likely quadrupeds. The dentition varied in different species, with some resembling that of the modern chimpanzee; some, the orangutan; and still others, the gorilla. The canine teeth were moderately tusklike but not as pronounced as those of modern apes. In summary, Dryopithecus is thought to be an ancestral form of the apes and possibly of the family Pongidae.
2.3.2 Ramapithecus. In 1932, G. Edward Lewis, a young Yale University graduate student, discovered in Haritalyanger, a cluster of villages some 100 miles (160 km.) north of New Delhi, India, a single fragment of an upper jaw of a so-called manlike ape. Subsequently, many similar specimens have been found in Miocene and Pliocene rocks in Turkey, central Europe, and East Africa. The specimens include Kenyapithecus, Graecopithecus, and Rudapithecus. The finds were mainly teeth and jaws. The dentition of Ramapithecus differs from that of Dryopithecus and approaches that of Australopithecus (see I.2.3.3), the most humanlike fossil primate. The smaller teeth, less prominent canines, thicker enamel, and other differences induced anthropologists to give this find the status of a new genus, Ramapithecus.
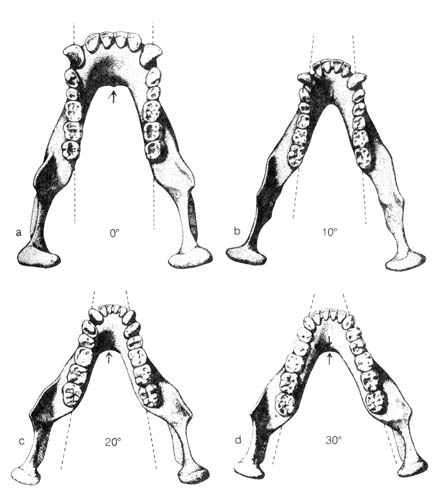
E. L. Simons has postulated that Dryopithecus gave rise to at least three genera between 10 and 15 million years ago. Two of them, Sivapithecus and Gigantopithecus, were apes with faces as large as those of the modern chimpanzee or gorilla. However, the third genus, Ramapithecus, had a distinctly small face. On the basis of this and the comparison of the rearward divergence of the tooth arcade in the fossil primates (Figure 2.15), he concluded that Ramapithecus is part of the family Hominidae that may have given rise to the later hominid Australopithecus. However, the interpretation of Australopithecus itself is now in jeopardy. The recent discovery of a possible Homo fossil living at the same time with an early Australopithecus has suggested that Australopithecus may not be in the direct lineage of the genus Homo at all. Therefore, this uncertainty, the scanty evidence for Ramapithecus (only jaws and teeth), and the absence of intermediate fossils during the 4-5 million years between the most recent find of Ramapithecus and the oldest Australopithecus fossil have made many skeptical of Simons's interpretation.
Figure 2.15. Four lower jaws show variations in the amount of rearward divergence of the tooth arcades in three fossil primates. For comparison, (a) is the mandible of a modern chimpanzee; its typically U-shaped dental arcade has parallel tooth rows; thus, the degree of divergence is zero. Next (b) is a reconstructed Dryopithecus mandible; the tooth rows show an angle of divergence (dotted lines) averaging some 10 degrees. Next (c) is a composite reconstruction of a Ramapithecus mandible. Its tooth rows, when preserved, show an angle of divergence averaging 20 degrees. Last (d) is a reconstructed Australopithecus mandible. Its typical angle of tooth-row divergence is 30 degrees. The tooth rows of later hominids show even greater angles of divergence. Arrows show differences in the two jaw-ridge buttresses known as the superior and the inferior torus. Modern apes possess a large, shelflike inferior torus; in Dryopithecus the superior torus was dominant. Both the ridges are developed in Ramapithecus and Australopithecus. Reprinted, with permission, from Simons, E. L. Ramapithecus. Sc. Am. 236 (May): 32-33; 1977.
Table 2.11. Sites yielding evidence of Australopithecus.* [100] 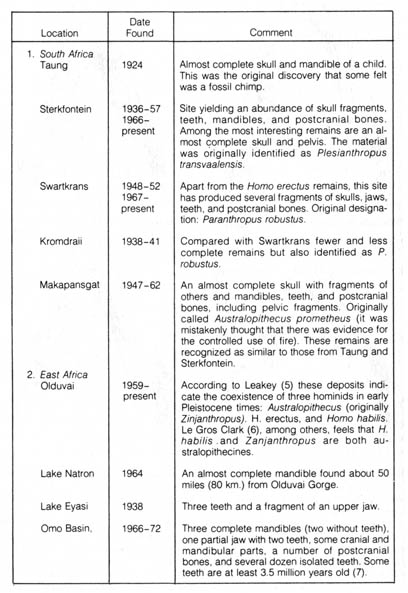
Table 2.11 (Continued) [101]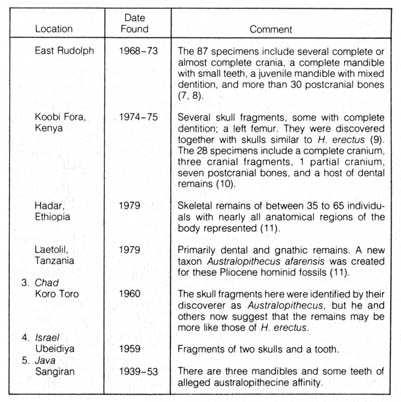
*NOTE: Adapted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970.
In surveying the fossil record pertaining to primate evolution,
one point becomes obvious and has an important implication: many of the
phylogenetic relationships postulated are based on very little fossil evidence.
Most of the more credible conclusions are obtained not by logical deduction
from the fossil finds but by imposing characteristics of modern representatives
onto the fossil record (1). Moreover, Mixter (3)
has pointed out that the origin of the distinct human feature of bipedalism
(being two-footed) is not substantiated by any fossil finds. Therefore,
although some primate paleontologists are dogmatic about their assertions
of human lineage, evidence is not conclusive.
2.3.3 Australopithecus. The first Australopithecus (4) find was discovered in 1924 in Taung, a village about 80 miles (128 km.) north of Kimberley, South Africa. It was the fragmentary remains of a child's skull, [102] five to seven years of age. The anatomist Raymond A. Dart judged it to be the remains of an extinct hominid, perhaps the missing link between humans and apes. He called it Australopithecus africanus or the South African Apeman.
The skull has a curious mixture of human and simian (apelike) features. It looks very much like that of an infant chimpanzee and is not consistent with the morphological pattern of later more positively identified hominid shills. However, Dart, and later Robert Broom, argued that the Taung child's age was inconsistent with its being a pongid because the brain's size was too large for its age. They believed that if it were a chimpanzee, five to seven years old, it should have begun to show adult characteristics such as crests and ridges of the skull and they noted that its dentition resembles the teeth of humans rather than those of chimpanzees or gorillas.
The evidence presented by one specimen is not convincing, and the difficulties of the interpretation of this find are complicated since most comparisons with other finds are based on adult characteristics. However, the interest sparked by the find has prompted a diligent search for more Australopithecus remains. At present, a considerable number of additional skeletons, more mature and some nearly complete, have been collected from many places, including South Africa, East Africa, Chad, Israel, and Java (Table 2.11).
However, the attempts to categorize the various finds of Australopithecus have met with some difficulties. First, the early descriptions and interpretations, reflecting the dominant opinions of that time, tended to maximize the differences not only between Australopithecus finds and modern humans, but among the Australopithecus themselves. Secondly, the scattered Australopithecus finds and scanty geological stratigraphic evidence in most cases, except for the Tuang child, made a temporal correlation of the different finds a difficult task. Adding to the complication, the finds were cut out during excavation and later investigated. They should have been categorized chronologically according to climatic and faunal change evidence found along with them.
The recently discovered Plio-Pleistocene vertebrate assemblages in eastern Africa have been employed in correlating different sites in East and South Africa with important implications of hominid evolution (12) involving species of Australopithecus. Australopithecus gracilis africanus, the smaller form, apparently occupied an earlier phase, and Australopithecus robustus, the larger form, was found in a later phase. Some authorities regard the A. gracilis africanus as hominid and classified it as Homo africanus and Homo habilis. [103]
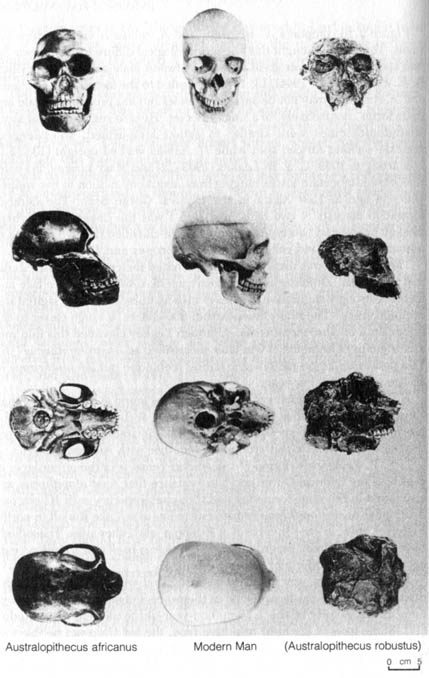
Figure 2.16 compares A. africanus and A. robustus with modern humans. Tobias (16) thought that the morphological differences between the two species were not as great as those between male and female gorillas. R. E. F. Leakey (b. 1944) (13, 14) also alluded to the fact that A. africanus and A. robustus may not be separate species but are probably female and male forms, respectively, of a single species. He also suggested that some australopithecines were similar to extant long-armed, short-legged, knuckle-walking African apes while H. habilis walked upright (13, 15).
Starting in 1959, L. S. B. Leakey (1903-72) and Mary Leakey, his wife, uncovered important fossils ranging from about one million to two million years in age in East Africa in the Olduvai Gorge beds. The Olduvai formation consists of two beds, and Bed I was the deeper layer where Leakey uncovered two types of remains. He classified the two types as of a hominid nature and called them Zinjanthropus and H. habilis (5).
The Zinjanthropus fossil unearthed included the cranium, tibia, fibula, and two teeth. Zinjanthropus is grossly apelike, with a massive face and features strongly suggestive of a heavy musculature, including sagittal and occipital crests. The brain size is within 600-700 cc, a range closer to the upper limit of Australopithecus. Although Leakey classified this find into a new genus of hominid, it was later reclassified as Australopithecus (16). The potassium-argon radiometric dating technique put Zinjanthropus at an age of 1.75 million years.
H. habilis is the remains of an infant and was found at a lower level than Zinjanthropus but still at Bed I. Although L. S. B. Leakey claimed it to be more humanlike than apelike, LeGros Clark (6), among many others, has discounted H. habilis and grouped it together with Australopithecus.
L. S. B. Leakey also found Homo erectus remains in the upper layer of Bed II of the Olduvai Gorge and had evidence that Australopithecus was also found in the same formation and thus contemporary with H. erectus. Additional Australopithecus remains were found at Lake Rudolf in northeastern Kenya, in the Omo basin of Ethiopia, and other sites. These finds have pushed the fossil record of Australopithecus back to 5.5 million years ago if one accepts the more liberal estimate.
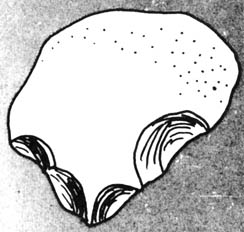
Australopithecus was found to be associated with toolmaking by
the discovery of stone flakes struck off in the process of shaping pebble
choppers in the vicinity of the 1.75-million-year-old fossil near Lake
Rudolf (17). Stone tools of the most primitive type,
the old Oldowan pebble tool (Figure 2.17), have been found with Australopithecus
fossils in three of the other five South African sites. Since wild chimpanzees
have been known to make crude tools (15, 17),
the toolmaking capacity of
Australopithecus was not entirely surprising.
However, a circular stone [104] structure, usually associated with shelter
building, found at the base of Bed I in the Olduvai Gorge contemporary
with H habilis is still not well explained (18). It is very unlikely
that the "artifact" was used for the manufacture of shelters since this
venture is believed to be a late human accomplishment occurring within
the last 100,000 years. However, the dissimilarity of this tool with the
other crude pebble tools makes it most perplexing.
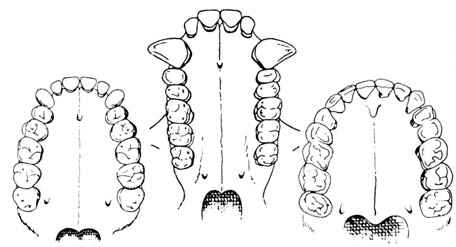
Figure 2.16 Comparison of four views of two adult australopithecine skulls with a skull of modern humans. Reprinted; with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippencott & Co.; 1970. [104] Figure 2.17. Oldowan pebble tool. Reprinted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970.
As indicated in Figure 2.16, Australopithecus, although resembling
the chimpanzee, differs significantly in its cranial capacity of 400-600
cc and in having a much larger body size (about four feet tall, slender
forms weighing around 50 pounds). The forehead is more rounded out, and
the eyebrow ridges are less prominent than in the chimpanzee. The jaws
also protrude less prominently than in apes. The dentition is quite similar
to that of modern humans (Figure 2.18). The occipital condyles, by which
the skull articulates with the spinal column, are set much farther forward
toward the ventral surface on the skull, suggesting a relatively upright
posture. The rest of the skeletal remains of Australopithecus generally
corroborates the evidence of the skull.
Although the factual findings of Australopithecus are almost universally accepted, their interpretations have been controversial. It is difficult to establish definitive relationships of Australopithecus with hominids. The temporal, spatial, formal, and contextual information from each locality of the finds has to be considered, and a satisfactory means must still be established to relate with much certainty the time of South African Australopithecus deposits to the glacial sequence in the northern latitudes. Moreover, in contrast with H. erectus and Homo neanderthalensis (see [106] I.2.3.4.b), which have some modern-day analogues or simulated analogues, Australopithecus has no modern-day counterpart. Although some scholars contend that these modern-day analogues have nothing to do with how a fossilized organism might have lived, others believe they would at least illuminate some ways of life of their ancestors. Therefore, the interpretation of the way Australopithecus lived is at best an educated guess.
Figure 2.18. Palate and upper teeth of an australopithecine; a gorilla; and a human; from left to right, respectively. Reprinted, with permission, from Clark, 10th ed. The history of primate. British Museum (Natural History), by permission of the Trustees of the British Museum (Natural History). 1970.
In the earlier days of its discovery anthropologists were reluctant
to accept Australopithecus into the lineage of human evolution.
After the successful campaign of Dart, Broom, and Clark with more finds,
most anthropologists placed Australopithecus in the direct line
of human descent. However, several recent findings have placed doubt on
this interpretation.
The first find was a living high-altitude Ethiopian baboon, Theropithecus
galada. It was found to have incisors and canines that are small relative
to those of extant African apes. Its cheek teeth are closely packed and
heavily worn. In addition, it also possesses many features shared by Ramapithecus
and Australopithecus, such as powerful masticatory muscles and a
short, deep face (19, 20). Since
the living baboon T. galada possesses some of the dental and facial
characteristics of Ramapithecus and
Australopithecus, a question
is raised about the validity of classifying the two fossils as hominids.
The baboon is living now, so presumably it is not directly related to man
phylogenetically. [107]
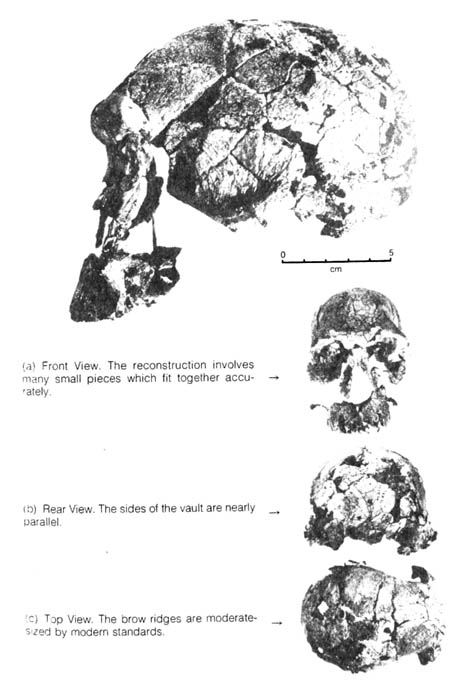
Figure 2.19. Cranium KNM-ER-1470, a new Homo from East Rudolph. Reprinted, with permission, from Professor Birdsell, Department of Anthropology, UCLA. From Birdsell, J. B. Human evolution. Chicago: Rand McNally; 1975. (a) Front View. The reconstruction involves many small pieces which fit together accurately. (b) Rear View. The sides of the vault are nearly parallel. (c) Top View. The brow ridges are moderate-sized by modern standards.
[108] The second find was a recent discovery by R. E. F. Leakey
at East Rudolf in northern Kenya of 150 fragments of a single skull (KNM-ER-1470)
in one stratum dated by the potassium-argon method to be 2.6 to 2.9 million
years old (21). The cranial capacity is 780-810 cc.
This is much larger than that of Australopithecus and very similar
to that of H. erectus (Figure 2.19). At the same time both A.
robustus and
A. africanus lineages were found at the
same stratum although some australopithecines less than one million years
old were also found. In addition, the Lake Rudolf skull is quite unlike,
and possibly more advanced than, the skull of H. erectus. Therefore,
Leaky classified it as
Homo sp. indet. (species indeterminate).
The antiquity of this Homo
species is also corroborated by the possible
remains of genus Homo
found in the Laetolil Beds near the Olduvai
Gorge in northern Tanzania. They were dated back to 3.59 to 3.77 million
years ago (21, 22). It is of interest
to note that the cranial capacity of KNM-ER-1470 is larger than that of
some primitive H. erectus though the two Homo species are
separated by almost two million years. The KNM-ER-1470 skull raises the
question that the earlier Australopithecus was contemporary with
Homo
sp. indet. and thus the australopithecines may have had nothing to
do with the evolution of the genus Homo.
![]()
References 2.3
1. Simons, E. L. Sc. Am. 236 (5):28; 1977.